
«
L'environnement n'est pas que co-présent; il est aussi co-organisateur.
[...]. L'environnement, loin de réduire son caractère co-organisateur,
l'accroît chez l'être vivant. Comme on le verra, l'environnement, devenu
éco-système, c'est-à -dire une machine spontanée née des interactions
entre les êtres vivants d'une même « niche », est beaucoup plus qu'une
réserve de nourriture, plus encore qu'une source de néguentropie où
l'être puise de l'organisation, de la complexité, de l'information, c'est
une des dimensions de la vie, aussi fondamentale que l'individualité,
la société, le cycle des reproductions.
Ainsi
s'impose l'idée clé : l'environnement est constitutif en permanence de
tous les êtres qui s'alimentent en lui; il coopère en permanence avec
leur organisation. Ces ĂŞtres et organisations sont donc en permanence
éco-dépendants. Mais, par un paradoxe qui est le propre de la relation
écologique, c'est dans cette dépendance que se tisse et se constitue
l'autonomie de ces ĂŞtres. De tels ĂŞtres ne peuvent construire et maintenir
leur existence, leur autonomie, leur individualité, leur originalité
que dans la relation écologique, c'est-à -dire dans et par la dépendance
à l'égard de leur environnement; d'où l'idée alpha de toute pensée
écologisée : l'indépendance d'un être vivant nécessite sa dépendance
à l'égard de son environnement. »
Edgar
Morin
(La
MĂ©thode, I). |
On donne le nom
de biosphère à la partie de notre planète
oĂą se rencontrent les organismes vivants. Cela
inclut la basse atmosphère, toute la région de la croûte terrestre la
plus proche de la surface (la pédosphère, épaisse de quelques mètres),
les océans et les mers (hydrosphère), ainsi que l'ensemble des organismes
présents dans cet espace.
On a proposé d'utiliser le terme d'écosphère
pour désigner la biosphère lorsqu'on envisage celle-ci non pas seulement
en tant que partie de la Terre abritant des organismes vivants, mais en
tant qu'écosystème (ou ensemble de tous les écosystèmes), c'est-à -dire
lorsqu'on s'intéresse à l'ensemble des processus en jeu lors des interactions
entre les différents organismes vivants et leur environnement. Cela implique
d'étendre l'étude à l'atmosphère
dans sa totalité, puisque sa composition en oxygène (dioxygène O2
et ozone O3)
découle entièrement de l'activité photosynthétique
des plantes, et que les rejets dûs aux activités humaines (gaz à effet
de serre, CFC et autres polluants) ont Ă©galement un impact global. Cela
revient aussi Ă ajouter la dimension temporelle et dynamique Ă l'Ă©tude
de cette biosphère et Ă l'envisager comme le rĂ©sultat de processus Ă
l'oeuvre depuis près de quatre milliards d'années, c'est-à -dire depuis
que les premiers organismes vivants sont apparus sur la Terre.
L'étude de la biosphère
et de l'écosphère conduit, par la définition même de son objet à considérer
deux ordres de phénomènes, ceux qui concernent
le vivant (facteurs biotiques) et ceux qui concernent tout ce qui n'est
pas vivant (facteurs abiotiques). Les interactions entre organismes vivants
(compétition, mutualisme, etc.), celles de caractère purement physique
et chimique (géophysique, géochimique), et celles entre les êtres vivants
et leur milieu, qui définissent à proprement parler le domaine d'étude
de l'écologie. Ces interactions se font via des échanges de matière
et d'Ă©nergie.
Les visages du vivant
L'Ă©volution du vivant.
La
chimie prébiotique.
L'existence d'organismes vivants
sur la Terre est attestée depuis , possiblement, 3,8 milliards d'années
(soit seulement 800 millions d'années après la formation de la planète).
Si l'on met Ă part l'Ă©nergie indispensable
à toute la chimie du vivant, c'est la présence d'eau liquide en abondance
qui a été la première condition à l'apparition de ces organismes. L'eau
liquide est un milieu qui permet à la fois la rencontre des éléments
chimiques qui peuvent se lier pour former des molécules de plus en plus
complexes. La molécule d'eau, est un agent réducteur (elle fournit un
électron lors de certaines réactions chimiques); en tant que dissolvant,
elle est en mesure de faciliter la rupture des liaisons entre molécules,
si bien qu'elle apparaît comme indispensable à toutes les transformations
chimiques opérées dans les organismes vivants.
L'autre élément essentiel à la chimie
du vivant est le carbone. Les caractéristiques de l'atome
de carbone (notamment sa tétravalence) le rendent apte à servir de ciment
aux édifices moléculaires éventuellement complexes (macromolécules)qui
sont à la base de tous les organismes vivants et des phénomènes qui
les caractérisent. Ces molécules sont formées d'un squelette d'atomes
de carbone auquel sont liés quantité d'autres éléments (oxygène, hydrogène,
azote, calcium, phosphore, soufre, potassium, etc.). La chimie du carbone
est par excellence la chimie du vivant et est ordinairement nommée, pour
cette raison, chimie organique.
Les molécules organiques qui ont précédé
les premiers organismes vivants se sont constituées à partir du carbone
disponible et de l'hydrogène, - celui-ci sans doute d'abord fourni par
les molécules de sulfure d'hydrogène H2S (agent
réducteur), puis par les molécules d'eau H2O. Certaines
de ces molécules (des acides
aminés parmi elles) ont pu aussi être synthétisées dans
l'espace et être apportés sur la Terre par des chutes de
météorites
ou des collisions avec des noyaux cométaires.
Le premier pas important sur le chemin qui menait aux organismes vivant
a été la formation des premières molécules capables de se répliquer.
Aujourd'hui, ce sont les molécules d'ADN qui, possédant
cette capacité, portent l'information nécessaire (information génétique)
pour que puisse se constituer un organisme vivant Ă partir d'une autre.
Dans un premier temps, ce rôle était dévolu à des molécules d'ARN
( Les
acides nucléiques), mais la grammaire générale de ce qui allait
être la reproduction des organismes et de la constitution des espèces
vivantes Ă©tait la mĂŞme. Les
acides nucléiques), mais la grammaire générale de ce qui allait
être la reproduction des organismes et de la constitution des espèces
vivantes Ă©tait la mĂŞme.
Descendance
et différence. Dès l'époque prébiotique, lorsqu'on parle de molécules
capables de se dupliquer, on n'entend pas qu'elles produisent des répliques
exactement identiques à elles-mêmes. Il peut exister des différences
entre la molécule fille et la molécule mère, dues à des erreurs aléatoires
dans la transmission de l'information génétique. Certaines de ces erreurs
ont pour conséquence une descendance plus ou moins apte à se reproduire
elle-mĂŞme (peut-ĂŞtre en fonction du milieu dans lequel vivent les organismes
concerné, ou pour d'autres raisons). C'est le principe de l'évolution,
qui va de pair avec la formation de divers chemins Ă©volutifs, certains
menant à l'apparition de nouvelles espèces, d'autres à la disparition
d'espèces existantes.
Un autre pas a été franchi quand des molécules
d'ARN capables de duplication se sont trouvées
enfermées dans des microsphères. De telles structures sont des membranes
repliées sur elles-mêmes (constituées de
lipides
et de protéines), qui ont pu se former spontanément,
si les conditions physiques et mécaniques étaient réunies. Une molécule
d'ARN capable de se répliquer et enfermée dans un espace délimité par
une membrane, cela commence Ă ressembler Ă une cellule
vivante. Mais il a fallu encore franchir quelques Ă©tapes pour que l'on
puisse parler d'organismes vivants.
Ces structures prébiotiques ont dû acquérir
notamment la capacité de ce maintenir telles quelles malgré les variations
possibles de leur environnement. Cela signifie, la capacité de maintenir
des conditions physiques et chimiques à peu près constantes à l'intérieur
de l'espace renfermé par la membrane. Cette capacité d'autorégulation
interne qui, après toutes les caractéristiques déjà évoquées, forme
le socle de la définition d'un organisme vivant, est appelée
homéostasie.
Un élément central de cette capacité de régulation interne est la possibilité
de gérer d'une manière ou d'une autre l'énergie qui provient (ou qui
éventuellement peut ne pas provenir pendant un certain temps) de l'extérieur.
Les toutes premières cellules
vivantes, notamment grâce à l'apparition chez elles, il y a environ
2,2 milliards d'années, de certaines structures spécialisées, appelées
organites,
ont dû acquérir cette capacité avant de perfectionner de diverses manières
leurs modes d'acquisition de l'énergie (comme la photosynthèse apparue
très précocement), de transformation et de stockage interne. Ces processus,
en mĂŞme temps que ceux qui concernent la transformation de matière Ă
l'intérieur d'un organisme vivant, en définissent le métabolisme.
-

| Stromatolithes
de Shark Bay (Australie Occidentale). - Les stromatolithes sont des
formations minérales que l'on rencontre dans les zones intertidales (
= bandes littorales délimitées par l'amplitude des marées). Elles sont
produites par des bactéries, notamment par des cyanobactéries. Ces micro-organismes
photosynthétiques détachent le carbone du CO2 et
le libèrent sous la forme de carbonates, qui, par précipitation, forment
les stromatolithes. Ces formations ont existé à peu près à tous les
moments de l'histoire de la Terre et se rencontrent sous des formes fossiles
en de nombreux lieux. Les stromatolithes ont connu leur âge d'or au Protérozoïque,
entre -2,2 milliards d'années et -570 millions d'années. Ils fournissent
certains des plus anciens indices de la présence d'organismes vivants
sur notre planète. Des stromatolithes ont ainsi été découverts au Groenland,
vieux de 3,7 milliards d'années. Photo : Paul Harrison
/ USGS. |
Procaryotes
et eucaryotes.
Dans les premières cellules, les molécules
renfermant l'information génétique étaient simplement contenues à l'intérieur
de la membrane
cellulaire, sans qu'un lieu bien délimité leur soit destiné,
comme c'est le cas encore aujourd'hui avec les bactéries
et les archées.
De tels organismes sont qualifiés de procaryotes. Mais l'évolution
de certaines cellules les a conduites à enfermer les molécules renfermant
l'information génétique dans une région spéciale, elle aussi circonscrite
par une membrane, un organite particulier, appelée le noyau cellulaire.
Les cellules dotées d'un tel noyau (et les organismes formés de telles
cellules) sont dites eucaryotes.
Unicelluaires
et pluricellulaires.
Les Eucaryotes sont les organismes qui
ont suivi les chemins évolutifs les plus variés. La plupart de ceux qui
sont restés constitués d'une seule cellule (organismes unicellulaires)
ont formé le gros du bataillon des Protistes dont les plus anciens remontent
à 1,8 milliards d'années; les autres ont constitué des ensembles de
cellules fonctionnant ensemble pour former des organismes pluricellulaires.
Depuis une découverte faite au Gabon en 2008,
on fait remonter les plus anciens organismes pluricellulaires Ă 2,2 milliards
d'années (Orosirien), mais ils pourraient avoir disparu assez rapidement.
Une autre souche, celle des algues rouges (Rhodophytes) remonte Ă 1, 2
milliards d'années (Sténien). Les ancêtres des organismes pluricellulaires
actuels (les animaux, les végétaux, les champignons) pourrait ne remonter
qu'à Ediacarien, il ya 600 millions d'années (Le
Passé de la Terre). Quoi qu'il en soit, les organismes pluricellulaires
possèdent des cellules spécialisées et en général des tissus différenciés,
chacun ayant ses propres caractéristiques et fonctions, et qui forment
des organes.
Autotrophes
et hérérotrophes.
Tous les organismes
sont constitués d'eau, de sels minéraux et de très divers composés
organiques. Ils puisent dans le milieu où ils vivent les ressources (lumière
du soleil, nutriments minĂ©raux et matière organique) nĂ©cessaires Ă
leur survie, Ă leur croissance Ă leur entretien et Ă leur reproduction.
La nutrition fournit
les matériaux nécessaires à la construction des molécules dont l'organisme
est constitué. Celles-ci sont dégradées ou transformées de telle sorte
que des échanges constants de matière avec le milieu extérieur sont
nécessaires. L'énergie, qu'elle soit apportée par la nutrition ou non,
est nécessaire dans la la plupart des processus métaboliques, en
particulier ceux responsables de la construction des macromolĂ©cules Ă
partir de composés plus petits; elle aussi se dégrade en permanence et
doit être renouvelée constamment.
Les organismes capables
de synthétiser leur propre nourriture (plus précisément, capables d'utiliser
du carbone inorganique comme source de carbone), et de d'obtenir par eux-mĂŞme
l'énergie dont ils ont besoin à partir de leur milieu sont appelées
autotrophes.
Ceux qui doivent se nourrir d'autres organismes pour acquérir leur énergie
sont appelés hétérotrophes (en grec,
trophè = nourriture,
aliments).
•
Les
autotrophes. - Un organisme autotrophe
(au carbone) est un organisme capable de synthétiser les molécules utilisées
pour la construction et le fonctionnement des ĂŞtres vivants en puisant
leur carbone dans des composés simples (dioxyde de carbone CO2,
dans le cas des organismes terrestres, acide carbonique, H2CO3
pour les autotrophes marins). Dans certains cas l'acquisition du carbone
inorganique se fait à partir de réactions chimiques inorganiques (chimiosynthèse),
mais le plus généralement en utilisant l'énergie lumineuse (photosynthèse),
qui est ensuite stockée dans des liaisons covalentes qui relient les atomes
de carbone entre eux pour former notamment des glucides
(sucres).
+ Les photoautotrophes,
tels que les plantes, les algues et les bactéries photosynthétiques,
exploitent l'Ă©nergie solaire (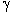 )
en la convertissant en énergie chimique d'abord sous forme d'adénosine
triphosphate (ATP), et d'une enzyme
qui intervient dans la photosynthèse, le nicotinamide adénine dinucléotide
phosphate (NADP). L'énergie stockée dans l'ATP est ensuite utilisée
pour synthétiser des molécules organiques telles que le glucose (CH2O).
Ainsi pourra-t-on résumer, au final, le processus de la photosynthèse,
par la réaction : )
en la convertissant en énergie chimique d'abord sous forme d'adénosine
triphosphate (ATP), et d'une enzyme
qui intervient dans la photosynthèse, le nicotinamide adénine dinucléotide
phosphate (NADP). L'énergie stockée dans l'ATP est ensuite utilisée
pour synthétiser des molécules organiques telles que le glucose (CH2O).
Ainsi pourra-t-on résumer, au final, le processus de la photosynthèse,
par la réaction :
CO2
+
H2O +  CH2O + O2
CH2O + O2
+ Les chimioautotrophes
sont principalement des bactéries que l'on rencontre là où la lumière
du soleil n'est pas disponible (grottes sombres, cheminées hydrothermales
au fond de l'océan). De nombreux chimioautotrophes des cheminées hydrothermales
utilisent comme source d'énergie chimique le sulfure d'hydrogène (H2S),
qu'elles libèrent. Les chimioautotrophes, comme les photoautotrophes,
synthétisent ensuite des molécules organiques comme le glucose pour y
stocker l'énergie requise par leur métabolisme, et qui sera aussi utilisable
par d'autres organismes.
• Les hétérotrophes.
- Un organisme hétérotrophe est un organisme incapable d'effectuer les
synthèses de ses constituantsdirectement à partir d'éléments
minéraux. Il doit puiser son énergie et ses nutriments sous forme de
matière organique en consommant d'autres organismes (autotrophes ou non).
En ce sens les hétérotrophes sont aussi chimiotrophes . Les animaux,
les champignons, certaines bactéries, les protozoaires sont hétérotrophes.
Certains organismes,
rangés parmi les autotrophes, sont, à certains moments de leur vie, ou
pour certaines de leurs parties aussi hétérotrophes. On parle alors de
mixotrophie.
La plupart des Protistes sont mixotrophes. Il en est de mĂŞme des plantes
vasculaires (Cormophytes), qui sont sont autotrophes pour ce qui concerne
leur partie aérienne, mais hétérotrophes par leurs racines.
Anaérobies,
aérobies.
Les premiers organismes Ă©taient
anaéorobies
: ils vivaient dans un environnement presque dépourvu d'oxygène (sa concentration
atmosphérique était inférieure à 2%). L'oxygène fabriqué par eux
(cyanobactéries
ou leurs ancêtres) au travers du processus de photosynthèse, était aussitôt
fixé par divers éléments et minéraux. Il était, par exemple, capté
par les
roches riches en fer (origine des formations
ferrifères rubanée du Sidérien, caratérisées par la magnétite, Fe3O4).
Quand cette captation n'a plus été possible, faute de récepteurs
disponibles (puits d'oxygène), cet élément - toxique pour
les organismes anaérobies - s'est accumulé aussi bien en dissolution
dans l'eau que dans l'atmosphère. Beaucoup des organismes existants alors
ont disparu (on parle de catastrophe de l'oxygène pour qualifier cet événement
qui a eu lieu il y a 2,4 milliards d'années).
Des Cyanobactéries ont cependant continué
à enrichir l'atmosphère en oxygène. Il a fallu du temps pour que se
développent des organismes aérobies, c'est-à -dire capables non
seulement de vivre dans un milieu riche en oxygène, mais d'utiliser cet
oxygène désormais disponible en abondance. L'utilisation de l'oxygène
par les processus métaboliques correspond à la respiration.
La
respiration aérobie se présente ainsi comme le processus inverse de la
photosynthèse.
On peut en schématiser les effets chimiques par
la réaction :
CH2O
+ O2
CO2 + H2O +
(La combustion lente
du glucose libère l'énergie renfermée dans les liaisons moléculaires
de celui-ci et produit du dioxyde de carbone et de l'eau).
La concentration de
l'oxygène dans l'atmosphère a crû à peu près régulièrement, pour
s'établir, il y a environ 500 milions d'années à des niveaux du même
ordre que ceux que l'on constate aujourd'hui. Chemin faisant l'oxygène
atmosphérique a permis un refroidissement du climat terrestre en faisant
diminuer la concentration atmosphérique du méthane (un gaz à effet de
serre).
A la fin du Protérozoïque,
l'oxygène, sous la forme d'ozone (trioxygène) a constitué dans la haute
atmosphère une couche capable de bloquer les rayonnements ultraviolets
du Soleil les plus nocifs pour les organismes vivants. Ceux-ci Ă©taient
jusque là confinés dans les océans ou du moins dans des milieux aquatiques;
l'augmentation de l'oxygène et la diminution du risque ultraviolet ont
facilité leur établissement sur la terre ferme et donné un grand élan
au développement des organismes pluricellulaire terrestres, d'abord végétaux
puis animaux. Ces conditions nouvelles ont ainsi préparé la grande époque
du vivant, marquée par une diversité de formes inédites, connue sous
le nom d'explosion cambrienne (vers
540-530 millions d'années avant le présent). Parallèlement, le phytoplancton
a aussi connu un essor considérable. Il réalise, à l'époque actuelle,
40 % de toute la photosynthèse.
-
| Evolution
de la concentration de l'oxygène (rouge) et du dioxyde de carbone
(noir) dans l'atmosphère pendant le Phanérozoïque (du Cambrien à aujourd'hui).
- L'atmosphère est restée réductrice jusqu'à il y a environ 600 milions
d'années. A partir de là , la concentration d'oxygène, après plusieurs
oscillations, avec un pic autour de 20% au moment de l'explosion du Cambrien,
est passée de 10% à un niveau proche de celui qu'elle a actuellement,
soit environ 21%. Au cours des derniers 400 millions d'années, elle n'a
oscillé que dans des limites relativement étroites autour de 24%. Ce
sont les organismes photosynthétiques qui en sont responsables. Le Carbonifère,
le Permien et le Crétacé
présentaient des concentrations d'oxygène élevées; le Trias
inférieur et moyen a été une période de plus faible teneur en oxygène;
du milieu du Trias supérieur au
Jurassique,
l'oxygène semble avoir fluctué mais est resté supérieur aux niveau
actuel. Depuis le Crétacé, le niveau d'oxygène a diminué.
Il
y a beaucoup moins de CO2 dans
l'atmosphère que d'oxygène, et les variations de la concentration de
dioxyde de carbone sont représentées sur le graphique à une échelle
très différente (en ppm = parties par million). On notera la grande chute
de la concentration de CO2 pendant
le Carbonifère, c'est-à -dire à l'âge d'or des cryptogames
vasculaires (des ptéridophytes, notamment).
Le carbone fixé ainsi par ces végétaux est l'actuel réservoir des combustibles
fossiles. Comme celle de l'oxygène, la concentration du dioxyde de carbone
a beaucoup oscillé. Le lissage de la courbe ne permet pas de figurer des
variations de courte durée, tels les pics à la fin du Permien ou du Crétacé
ou encore la montée vertigineuse du CO2 atmosphérique
(de 278 ppm à 420 ppm) depuis le début de l'ère industrielle.
(Sources
: K. Cantner, AGI et Foster et al.). |
Le vivant en ses
royaumes.
Dans les classifications
habituellement utilisées du monde vivant, on
distingue six règnes (archées, bactéries, protistes, végétaux, champignons
et animaux), entre lesquels se distribuent toutes les espèces actuelles.
Les
Archées et les Bactéries.
Les archées et les bactéries
sont des organismes procaryotes et presque tous unicellulaires. Ils sont
presque tous microscopiques (les plus grosses bactéries peuvent cependant
atteindre presque un millimètre de longueur). Ces deux groupes, dont l'évolution
séparée a commencé à peu près 200 millions d'années après l'apparition
des premiers organismes vivants, se distinguent par leur biochimie. On
les rencontre principalement dans les zones humides.
Les
Protistes.
Les protistes sont généralement des
organismes unicellulaires. Certains, à l'image du phytoplancton qui dérive
dans les océans et les
lacs,
sont photosynthétiques, comme les plantes et
utilisent donc l'Ă©nergie du soleil pour fabriquer de la nourriture. D'autres,
à l'image des amibes, du blob (Physarum polycephalum) ou des paramécies,
ont davantage d'affinités avec les animaux, on les range parmi les protozoaires.
Les protistes vivent dans l'eau ou dans les milieux humides.
Les
Végétaux.
Les végétaux
sont des organismes pluricellulaires vivant sur la terre et parfois dans
l'eau. Certains sont très simples (mousses,
fougères,
ou les algues, ces dernières parfois rangées
parmi les protistes), d'autres plus complexes, Ă l'image des spermatophytes
(plantes à graines). Tous ont pour caractéristique de pouvoir utiliser
presque toute l'énergie qu'ils reçoivent de la lumière solaire pour
opérer la photosynthèse.
Les premières algues
marines remontent au Cambrien. Les premières plantes vasculaires
apparaissent Ă l'Ordovicien,
elle commencent Ă envahir la terre, pour prendre une grande extension
au Carbonifère (grandes forêts de fougères
arborescentes et de lycopodes se développent,
premiers
conifères), qui correspond aussi
à l'apparition des premiers pollens. Les premières
forêts de gymnospermes (conifères, etc.)
remontent au Triassique. Les plantes Ă graines deviennent dominantes Ă
partir du Crétacé.
Les
Champignons.
Les champignons,
comme les végétaux parmi lequels on les rangeait autrefois, sont généralement
fixés au sol ou sur un support solide, et possèdent des racines
(leur système racinaire est souvent bien plus vaste que leur partie visible,
et peut atteindre plusieurs kilomètres carrés pour certaines espèces).
Mais, dépourvus de la capacité de photosynthèse, il sont rangés aujourd'hui
dans un règne distinct. Ils se nourrisent de matière organique (de plantes
mortes, en particulier). Les champignons sont généralement pluricellulaires,
mais quelques-uns, les levures (saccharomycètes), sont microscopiques
et unicellulaires. Les plus anciens champignons remontent au Silurien
ou Ă la fin de l'Ordovicien. (Certains champignons
inférieurs, généralement photosynthétiques, peuvent aussi être détachés
des champignons proprement dits, et constituer un règne supplémentaire,
celui des chromistes).
Les
Animaux.
Certains animaux
ne mesurent pas plus d'un millimètre de longueur, d'autres, comme la Baleine
bleue (Balaenoptera musculus), peuvent atteindre les 30 m. Les animaux,
qui vivent aussi bien sur la terre que dans les milieux aquatiques, se
distinguent des plantes par leur incapacité à synthétiser leurs aliments
(à commencer par les sucres) à partir de composés chimiques bruts. Ils
survivent en mangeant d'autres ĂŞtres vivants, ou des choses qui Ă©taient
autrefois vivantes (des plantes et des champignons, voire des protistes
(plancton), ou d'autres animaux). Si l'on excepte
queques animaux aquatiques qui restent pratiquement toute leur vie arrimés
Ă un support fixe (les mouvements de l'eau leur apportant la nourriture
nécessaire), la plupart des animaux sont capables de locomotion, soit
grâce à des organes spécialisés (nageoires,
ailes,
pattes),
soit par les mouvements de leur corps tout entier. La plupart des animaux
ont une
tête sur laquelle sont placés plusieurs
organes sensoriels (yeux, narines, oreilles),
qui fournissent une aide pour les déplacements et l'identification de
la nourriture. Il est commode de distinguer parmi les animaux deux groupes
:
+ Les Invertébrés, qui
représentent plus de 95 % des espèces animales actuellement connues.
Ils ne possèdent pas de colonne vertébrale et
de squelette interne articulé. Parmi eux : les vers, les insectes,
les mollusques,
les cnidaires (méduses, coraux), etc.
+ Les Vertébrés,
très minoritaires, qui possèdent, comme leur nom l'indique, une colonne
vertébrale et un squelette interne. Parmi
eux : les poissons, les amphibiens,
les reptiles, les oiseaux
et les mammifères.
Les premiers animaux
étaient des invertébrés marins au corps mou, et sont apparus il y a
environ un milliard d'années. L'explosion cambrienne a profité à l'essor
des animaux (métazoaires) : spongiaires,
arthropodes,
vertébrés. La plupart des invertébrés marins , à commencer par les
céphalopodes,
remontent à l'Ordovicien. A cette époque, les arthropodes, les céphalopodes
et les autres mollusques commencent Ă dominer
les mers, et sont rejoints par les poissons, de plus en plus présents
Ă partir du Silurien. Au DĂ©vonien, les insectes
et les premiers tétrapodes apparaissent. Les amphibiens, les reptiles
et les insectes formeront les principales classes d'animaux jusqu'Ă l'apparition
des dinosaures (Triassique) et des mammifères
(Jurassique). La disparition des Dinosaures lors de la transition Crétacé-Tertiaire,
il y a 65 millions d'années, permettra aux Mammifères de prendre une
place de plus en plus importante.
Nombre d'espèces
répertoriées et nombre estimé d'espèces
| Domaine |
Règne |
Espèces
répertoriées |
Nombre
d'espèces
estimé |
| Procaryotes |
Archées |
175 |
? |
| Bactéries |
10
000 |
? |
| Eucaryotes |
Protistes |
80
000 |
600
000 |
| Champignons |
72
000 |
1
500 000 |
| Plantes |
270
000 |
320
000 |
| Animaux |
|
| Arthropodes |
| |
Mandibulates
Crustacés
Chélicérates |
Mollusques
NĂ©matodes
| Vertébrés |
| |
Poissons
Amphibiens
Reptiles
Oiseaux
Mammifères |
|
|
| - |
|
70 000
25
000
| 52
500 |
| |
25
000
4950
8002
9750
4630 |
|
|
| 1
320 000 |
|
| - |
|
8
000 000
150 0000
750
000 |
200 000
400
000
|
|
| Total |
1
750 000 |
14
000 000 |
Sources
: UNEP / World Atlas of biodiversity, 2003.
Les grandes divisions
de la biosphère et de l'écosphère
Les organismes vivants
se répartissent dans la biosphère en fonction des conditions qui leur
sont offertes, notamment selon la disponibilité de l'eau, de l'énergie
et de la nourriture.
Les
Ă©cozones.
Les Ă©cozones
sont les plus grandes divisions biogéographiques de la surface de la Terre,
et se distinguent les unes des autres par des distributions caractéristiques
de la flore et des animaux qui y vivent. Elles correspondent à des régions
où les plantes et les animaux se sont développés dans un isolement relatif
pendant de longues périodes, du fait de divers facteurs géographiques
(océans, chaînes de montagnes,
déserts,
etc), qui ont constitué des obstacles à la migration des plantes ou des
animaux. Une écozone est donc définie selon les relations de parenté
entre les espèces vivantes qu'on y rencontre. Ces relations atant elles-mêmes
corrélées avec une histoire géographique et géologique commune. On
en compte huit.
•
La
région paléarctique comprend l'Europe, l'Afrique
septentrionale, le nord et le centre de la PĂ©ninsule
arabique, ainsi que l'Asie au nord de l'Himalaya.
On y range l'Islande, les îles de l'Arctique
eurasiatique et l'archipel japonais (sauf l'archipel
Ryukyu). Cela représente près de 55 millions de kilomètres carrés.
• La région
néarctique comprend la plus grande partie de l'Amérique
du Nord (dont le Groenland), au jusqu'au
Sud du Mexique (Hauts plateaux).
• La région
néotropicale, commence au Sud du Mexique (Terres chaudes) et s'étend
jusqu'au Sud de l'Amérique du Sud (régions
tropicales et régions tempérées). Elle comprend aussi les Antilles.
• La région
afrotropicale ou Ă©thiopique comprend toute l'Afrique
subsaharienne, Madagascar et les îles voisinnes
le sud de la PĂ©ninsule arabique, ainsi que le Sud de l'Iran
et du Pakistan.
• La région
indomalaise ou orientale s'Ă©tend sur la plus grande partie
de l'Asie du Sud, du Sud du Pakistan au Sud de la Chine
(provinces du Guangxi, Yunnan et Hainan) et jusqu'aux Philippines,
Ă Taiwan, aux Ă®les Ryukyu , Ă
Bornéo
et à l'archipel indonésien (à l'Ouest de
Lombok).
• La région
australasienne recouvre à peu près ce que l'on appelle l'Australasie.
Elle est séparée de la région indo-malaise par la Ligne
de Wallace (entre
Bali et Lombok).
Elle comprend l'est de l'Indonésie, l'Australie,
la Nouvelle-Guinée. On y ajoute parfois
plusieurs groupes d'îles du Pacifique,
que l'on détache de la région océanienne : le Vanuatu,
les îles salomon, la Nouvelle-Zélande,
La Nouvelle Calédonie, etc.
• La région
océanienne comprend les îles du Pacifique de Polynésie,
de Micronésie et de Mélanésie
(sauf éventuellement les îles précédemment nommées).
• La région
antarctique comprend l'Antarctide et
divers groupes du Sud de l'océan Indien
(îles Crozet, Kerguelen,
etc) et de l'océan Atlantique (îles
Shetland du Sud et GĂ©orgie du Sud,
etc.).
Les limites entre les
écozones ne sont pas toujours bien définies. A l'image de la ligne Wallace
(L'Océanie),
ces limites sont moins des lignes que des bandes plus ou moins Ă©tendues,
qui définissent des écoclines, autrement dit des gradations entre
deux zones. A l'intérieur de ces zones de transition entre deux faunes
et flores caractéristiques des écozones concernées, différentes lignes
pouvent souvent être tracées en fonction des groupes de plantes ou d'animaux
considérés.
Les
écorégions.
Les écorégions
sont des zones géographiques étendues auxquelles les divers caractères
(climat, géomorphologie, hydrologie, faune et flore, éventuellement les
caractères réunis conférés par l'activité humaine) confèrent une
écologie particulière. L'Amazonie, les Everglades
en Floride, la forĂŞt du Sri
Lanka, la savane soudanienne, la
forêt mixte de l'Europe méditerranéenne, par
exemple, sont des écorégions.
Les
biomes.
La notion de biome
se rapproche de celle d'écorégion, mais possède un caractère plus générique
(ex. : la taïga sibérienne et la taïga canadienne sont des écorégions;
la taïga tout court est un biome). Les biomes sont des zones de la biosphère
définies par le climat et la géographie et présentant des conditions
climatiques similaires. Ils sont définis sur la base de facteurs tels
que les structures végétales (arbres, arbustes et herbes), les types
de feuilles (feuilles larges et aiguilles), l'espacement des plantes (forĂŞt,
bois, savane) et le climat. Contrairement aux écorégions et aux écozones,
les biomes ne sont pas définis par des similitudes génétiques, taxonomiques
ou historiques.
On distingue les
biomes
terrestres (répartis en trois groupes principaux : forêts, formations
herbacées, déserts), les biomes aquatiques et les biomes mixtes.
Ces-derniers sont définis à partir des conditions particulières nées
de la rencontre de zones terrestres et de zones aquatiques (eaux peu profondes,
plages, bords de rivières, etc.) .
Les biomes terrestres
reçoivent leur nom en fonction de leur formation végétale typique et
pleinement développée ou d'une caractéristique marquante de son environnement
abiotique. Les caractéristiques du macroclimat existant sont souvent utilisées
à cet effet. Les principaux biomes terrestres sont signalés en gras dans
le tableau ci-dessous. Ajoutons, que les biomes terrestres peuvent ĂŞtre
divisées en entités plus petites, les paysages, possédant une
physionomie propre.
L'oecumène.
- Ce mot désigne l'ensemble des terres habitées en permanence et transformées
directement par l'espèce humaine. Elle est formée d'un ensemble de biomes,
dits anthropogéniques, dans lesquels on distingue divers types de paysages
(paysages urbains, paysages ruraux, etc.). On estime que l'oecumène recouvre
aujourd'hui la moitié de la surface terrestre.
Les biomes aquatiques
sont le plus souvent été définis à partir de caractères purement géographiques;
division en biomes continentaux (lacs, cours d'eau) et en biomes océaniques,
pour lesquels (comme dans le cas de certains lacs) la notion de profondeur
est prise en compte, car elle commande notamment à l'accès à l'énergie
solaire. Les régions supérieures sont atteintes par la lumière (on parle
de zone photique) et permettent la photosynthèse. Dans les régions
inférieures, sombres, (zone aphotique), l'accès à l'énergie (cela signifie
aussi Ă la nourriture) doit se faire selon des modes particuliers.
-
Les principaux
biomes
| ForĂŞts |
ForĂŞts subarctiques |
Forêts de conifères
(taĂŻga) |
| ForĂŞts
tempérées |
Forêts tempérées
de conifères tempérées |
| ForĂŞts humides
(feuillus ou conifères) |
| ForĂŞts de feuillus |
| Forêts mixtes (espèces
décidues et sempervirentes mélangées) : forêts méditerranéennes
ou sclérophylles
(bois, broussailles, maquis); forĂŞts de feuillus
et de conifères |
ForĂŞts
tropicales
et subtropicales |
ForĂŞts pluviales
Ă©quatoriales
ForĂŞts
de nuage |
| ForĂŞts de
mousson
(sèches par périodes) |
| ForĂŞts tropophiles
(forêts tropicales sèches) |
| FĂ´rets tropicales
de conifères (forêts de pins, forêts de cyprès). |
| Formations
herbacées |
Savanes |
Savanes herbeuses
(Llanos, en Amérique du Sud). |
| Savanes arbustives |
| Savanes Ă©pineuses,
broussailles |
| Landes,
bruyères |
| Prairies |
| Steppes |
Steppes proprement
dites, pampas, veldt, etc. selon les régions |
| Toundras |
Toundras arctiques |
| Toundras alpines
(hauts plateaux) |
| DĂ©serts |
DĂ©serts
froids |
Glaces : glaciers,
inlandsis(des
microorganismes ont été découverts en 2014, vivant à 800 m de profondeur
sous les glaces de l'Antarctide). |
| Rochers |
| DĂ©serts
chauds |
DĂ©sert de sable
(erg, au Sahara) |
| DĂ©sert de roches
(reg, hammada, sérir, au Sahara) |
| Oasis |
| Biomes
anthropogéniques |
Terres agricoles |
Terres labourées,
jardins, vergers, vignobles, rizières, pâturages,
etc. |
| Zones urbanisées |
Villes, habitats
humains divers, bordures des voies de communication (routes, voies ferrées). |
Biomes
aquatiques
(hydrobiosphère) |
Eaux
continentales
(environ
1,8 % de la surface du globe) |
Lacs,
Ă©tangs |
| Cours
d'eau |
| Deltas |
Eaux
océaniques
(environ
70 % de la surface du globe) |
Océans ouverts
(milieu pélagique, près de la surface; milieu démersal
'Ă des profondeurs plus grandes) |
| Grandes profondeurs,
fosses abyssales |
| Plateaux continentaux,
mers fermées, zones néritiques ou sublittorales |
| Zones d'algues flottantes |
| Banquises |
| Biomes
mixtes |
Zones
humides |
Marais,
prairies, savanes et forêts inondées ou inondables, tourbières |
|
Milieux
littoraux
Zones
intertidales (espaces entre les marées
hautes et basses) : vasières, plages, estuaires
Mangroves |
| Zones
ripariennes (bordures de rivières et
de lacs) : forĂŞts
galeries, etc. |
| RĂ©cifs
coralliens |
| Zones
benthiques |
Fonds océaniques,
fonds lacustres; limons |
| ForĂŞts d'algues,
herbiers
marins |
| Events océaniques
: cheminées hydrothermales, évents froids, sources sous-marines |
| Montagnes
: Ă©tagement de plusieurs biomes selon l'altitude.
Une grande variété d'écosystèmes, représentatifs de différents biomes
peuvent ainsi se rencontrer dans des espaces très réduits. |
Biomes des
milieux extrĂŞmes |
Milieux
dans lesquels les conditions physiques et chimiques "extrĂŞmes" : sources
hydrothermales, pergélisol,
grottes, l'intérieur des roches et des coraux, centrales nucléaires,
etc. |
Les écosystèmes
Les systèmes dynamiques.
Au sens le plus
large, un système est un ensemble dont les éléments entretiennent entre
eux des relations qui les rendent interdépendants et confèrent ainsi
une structure à cet ensemble. Cette définition conduit à s'interroger
sur ce que devient le système au fil du temps, du fait même de l'existence
de telles relations.
Lorsque ces relations
induisent des modifications dans le système on parle d'interactions. Un
système concret évoluant ainsi en fonction des interactions entre ces
éléments est appelé système dynamique. Un tel système peut
conserver ses caractéristiques (les paramètres qui le définissent)
dans la durée, se transformer pour se placer dans un état différent,
ou, à l'inverse, perdre complètement sa structure pour se disloquer.
La grandeur physique
utilisée pour caractériser une transformation et sa possibilité à l'intérieur
d'un système est l'énergie : on envisage les trasformations à l'intérieur
d'un système par le biais des échanges d'énergie entre les différentes
composantes du système lors de leurs interactions. Dans un système fermé,
l'énergie (et la matière) dont use le système est limitée par la définition
même du système. Au fil des transformations, elle perd de son efficacité
(deuxième principe de la thermodynamique)
et, au final, aucune modification à l'intérieur du système n'est plus
possible. A l'inverse, dans le cas d'un système ouvert, celui-ci
est alimenté de l'extérieur en énergie (et éventuellement en matière).
L'évolution du système se comprend alors en termes de flux d'énergie
: l'énergie entrante permet les transformations à l'intérieur du système,
et l'inévitable dégradation de l'énergie (qui la rend de moins en moins
efficace) est compensée par le maintien du flux entrant. Lorsque, sous
l'effet d'un flux entrant régulier un système est en mesure d'actionner
ses différents composants afin de maintenir son équilibre, il est dit
dans un Ă©tat stationnaire.
Quand on raisonne
en termes de flux, deux types de processus, appelés boucles de rétroaction,
interviennent dans l'évolution, l'équilibre ou non, d'un système, c'est-à -dire
dans sa régulation :
•
La rétroaction positive correspond à la situation où les effets
d'un processus agissent de telle sorte que ce processus soit renforcé
(effet boule de neige). Ce type de rétroaction est nécessaire pour activer
ou stimuler une transformation, une interaction, mais il aboutirait Ă
un emballement incontrolable si un deuxième type de processus n'intervenait
pas :
• La rétroaction
négative correspond à la situation où les effets d'un processus
font que le moteur de celui-ci est inhibé (effet de soupape).
Ces deux processus combinés
définissent la manière dont le système se régule. La capacité d'un
système à se réguler de telle sorte que ces conditions internes restent
confinées dans des limites généralement étroites est appelé homéostasie(homéostasie
dynamique).
La régulation d'un
système fait aussi appel à une autre notion, celle de facteur limitant
:
•
Un facteur limitant, aussi appelé limite de tolérance,
désigne une ressource, un paramètre ou une condition d'un système qui
limite l'évolution de tel ou tel processus à l'intérieur du système.
Cela peut être l'élément sur lequel agit la rétroaction négative,
mais la notion acquiert toute sa pertience lorsqu'elle s'applique Ă une
ressource rare requise par le système considéré.
Les systèmes sont exposés
à de multiples perturbations extérieures. Deux paramètres peuvent servir
à mesurer l'évolution d'un système face aux perturbations : la résistance
et la résilience.
•
La résistance est la capacité d'un système à maintenir son équilibre
malgré les perturbations.
• La résilience
est fonction de la vitesse à laquelle un système retrouve son équilibre
après avoir été perturbé. Une perturbation peut modifier un système
au point qu'il perde entièrement toute résilience. Ce processus peut
conduire à la destruction complète ou à l'altération irréversible
du système.
Les systèmes dynamiques
sont d'autant plus aptes Ă maintenir leur Ă©quilibre qu'ils sont complexes,
car cela multiplie d'autant les boucles de rétroaction qui gouvernent
cet Ă©quilibre.
Les composantes
des écosystèmes.
Un écosystème
est un système dynamique ouvert, défini dans un espace donné, qui possède
une composante vivante (biotique) et une composante non-vivante (abiotique).
L'écosphère est l'écosystème qui renferme tous les autres écosystèmes,
à commencer par les différents biomes, qui peuvent eux-mêmes se définir
comme composés d'écosystèmes de tailles très diverses.
On
rend parfois synonymes les mots écosystème et environnement,
mais on doit noter qu'un environnement (qui n'a pas nécessairement de
composante biotique) doit environner « quelque chose » : on devrait ainsi
parler de l'environnement d'une communauté biologique, d'un organisme
vivant, etc. C'est l'environnement plus le « quelque chose » qu'il
environne qui pourra donc éventuellement être assimilé à un écosystème;
quant à l'écosystème, qui est un système ouvert, il a, lui, à coup
sûr un environnement.
Les perturbations extérieures
auxquelles un écosystème peut être exposé sont, par exemple, les effets
du changement
climatique ou simplement des aléas météorologiques
(variations des températures et des précipitations, sécheresses, incendies,
inondations, etc.), les effets de l'activité humaine (pollution, déforestation
et autres formes d'anthropisation des sols, introduction d'espèces invasives,
etc.).
On distingue dans
un écosystème :
•
La
biocĂ©nose ou communautĂ© (ce dernier terme tendant aujourd'hui Ă
supplanter le précédent), renferme la totalité de organismes vivants
de l'écosystème. Ces organismes interagissent entre eux et induisent
la structure de l'écosystème.
+ Le
biote. - La biocénose (en grec, koinós = en commun)
ou communauté se distingue du biote en ce sens que la biocénose
désigne des éléments vivants en interaction, et partant composants d'un
système, alors que le mot biote n'a pas ce caractère dynamique : un biote
est seulement une collection d'êtres vivants dans un espace déterminé.
Pour définir un biote, il suffit d'en énumérer les espèces présentes;
pour définir une biocénose, il faut de surcroît préciser qu'elle en
est l'organisation, la diversité, la richesse spécifique.
+ La biomasse.
- Le mot biomasse désigne en premier lieu la masse (ou la masse par unité
de surface) des organismes vivants présents vivants dans une zone ou un
écosystème donné, à un moment donné. Le mot peut s'appliquer à la
masse de la communauté toute entière ou seulement à celle de certaines
des espèces. D'autres usages du terme se rencontrent : il peut aussi
se référer seulement à la masse de matière organique renfermée par
les organismes concernés (on ignore alors la masse de l'eau que referment
ces organismes), Ă la masse de carbone, etc. Dans ces derniers cas,
le terme de biomasse fait ordinairement référence à la quantité de
matière organique susceptible d'être utilisée comme source d'énergie
(bioénergie).
On définit à l'intérieur
d'une communauté biologique diverses composantes-:
+ Selon
les règnes qui sont considérés, on distinguera : la
phytocénose
(la flore), la zoocénose (la faune), la mycocénose
(les champignons), la microcénose (les microorganismes), etc.
+ En terme d'espèces
: les organismes d'une même espèce à l'intérieur d'une communauté
forment une population.
• Le biotope
(du grec topos = lieu) est la composante de l'écosystème
caractérisée par des facteurs abiotiques. Un biotope est ainsi une une
zone limitée en taille où vivent des organismes de même espèce ou d'espèces
différentes et où règnent des conditions physiques et chimiques définies
(relativement uniformes dans l'espace et stables dans le temps), telles
que la température, l'humidité, le vent, la lumière, le pH, etc. Un
biotope peut ĂŞtre une souche d'arbre, la forĂŞt dans laquelle se trouve
cette souche, un étang, un verger, une tourbière, une vallée, etc.
+ L'habitat.
- En général, le terme d'habitat (du latin habitare = vivre)
a un sens similaire à celui de biotope, mais alors que le biotope définit
le lieu de vie d'une communauté biologique, l'habitat se réfère à une
espèce particulière, et correspond au lieu de vie d'une population. Ainsi
entendu, l'habitat comprend l'espace que les individus d'une espèce parcourent
pour combler leurs besoins (se nourrir, se cacher, se reposer, se reproduire,
etc.) et peut aussi comprendre leur territoire de migration. Cependant,
certains auteurs anglo-saxons identifient habitat et biotope, quand d'autres
définissent un habitat comme une zone déterminée d'un biotope.
•
Niche
écologique. - Toutes les espèces ont une niche écologique dans l'écosystème.
La notion de niche écologique sert à caractériser la manière dont ces
espèces acquièrent les ressources qui leurs sont nécessaires et comment
elles interagissent avec les autres espèces de la communauté.
Il existe divers manières d'aborder une niche écologique. Par exemple,
a) selon l'habitat dans lequel vit une espèce et les adaptations
comportementales qui l'accompagnent (niche grinnellienne), b) selon la
manière dont une espèce ne se contente pas de se développer dans un
environnement, mais peut aussi modifier l'environnement et son comportement
au cours de sa croissance (niche eltonienne), ou encore c) en fonction
de divers paramètres statistiques et mathématiques qui définissent la
manière dont une espèce coexiste avec les autres composantes de la communauté
(niche hutchisonienne).
Les relations trophiques.
La
chaîne trophique.
La circulation de
l'énergie et de la matière au sein d'une biocénose est commandée par
les relations alimentaires qu'y entretiennent ses différentes composantes
(plantes, herbivores, carnivores,
décomposeurs),
et se décrit au travers de la notion de chaîne trophique ( = chaîne
alimentaire), qui est une représentation simplifiée des interactions
entre les différents organismes dans un écosystème. Une telle chaîne
illustre la façon dont les organismes se nourrissent les uns des autres
et comment l'énergie est transférée d'un organisme à un autre. Dans
une chaîne alimentaire, on distingue en particulier, les producteurs,
les consommateurs et les décomposeurs :
• Les
producteurs (plantes vertes, algues et certaines bactéries photosynthétiques),
sont des organismes capables de produire leur propre nourriture par photosynthèse.
Ils convertissent l'énergie lumineuse en énergie chimique, stockée sous
forme de matière organique (sucres, glucides, etc.).
• Les consommateurs
sont des organismes qui se nourrissent d'autres organismes pour obtenir
de l'énergie et des nutriments. Ils peuvent être classés en plusieurs
catégories :
+ Les
herbivores sont des consommateurs primaires qui se nourrissent de plantes
et d'autres producteurs.
+ Les carnivores
sont des consommateurs secondaires ou tertiaires qui se nourrissent principalement
de viande et d'autres animaux.
+ Les omnivores
sont des consommateurs qui se nourrissent Ă la fois de plantes et d'animaux.
• Les
décomposeurs sont des organismes qui décomposent la matière organique
morte et la décomposent en éléments nutritifs simples. Ils jouent un
rôle essentiel dans le recyclage des éléments nutritifs dans l'écosystème
en libérant les nutriments stockés dans les débris et les organismes
morts.
Les chaînes trophiques
peuvent être simples ou complexes en fonction de la diversité des espèces
dans l'écosystème et des nombreuses interactions qui s'y produisent.
Le
réseau trophique.
Les chaînes alimentaires
ne décrivent pas de façon complète les flux d'énergie et de matière
dans un écosystème. Même quand tous les organismes sont regroupés dans
les niveaux trophiques appropriés, on constate que certaines espèces
peuvent se nourrir Ă partir de plusieurs niveaux trophiques. On a dĂ©jĂ
signalé le cas des décomposeurs, on peut aussi penser aux omnivores qui
peuvent se nourir aussi bien de carnivores, que d'herbivores ou de plantes.
De plus, dans un écosystème, il est rare que l'on n'observe qu'une unique
chaîne alimentaire. Il en existe ordinairement plusieurs, liées entre
elles pour former une structure complexe. Cela amène à introduire la
notion de réseau trophique. Graphiquement, on exprime les relations
trophiques d'une communauté dans un tel réseau avec des flèches qui
relient les espèces en fonction de qui mange qui.
-
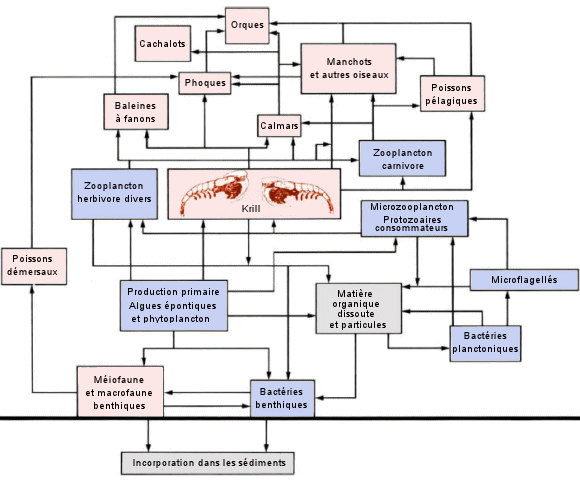
| Le
réseau trophique dans l'océan Antarctique. - Les réseaux alimentaires
marins polaires ont tendance Ă ĂŞtre plus simples que ceux des autres
écosystèmes (cela les rend particulièrement fragiles). Dans l'océan
Austral (ainsi d'ailleurs que dans l'océan
Arctique), il existe deux types fondamentaux de producteurs primaires
: le phytoplancton pélagique et les algues qui vivent dans la glace (algues
épontiques). Celles-ci sont généralement des espèces benthiques adaptées
aux faibles intensités lumineuses régnant sous la surface de la glace.
L'abondant krill (Euphausia superba) constitue le pivot du réseau trophique
de l'Antarctique car il en est le principal herbivore, et il constitue
aussi une source de nourriture importante pour plusieurs espèces de zooplancton
carnivore, de poissons pélagiques et de calmars, ainsi que pour les baleines
Ă fanons, les phoques et les oiseaux de mer qui se nourrissent de plancton.
(D'après
C. Lalli et T. Parsons, Biological oceanography, an introduction,
1993). |
Comme les réseaux
trophiques peuvent être très compliqués, on peut les simplifer de diverses
manières : par exemple en regroupant les espèces ayant des relations
trophiques similaires dans une communauté donnée en grands groupes fonctionnels,
ou encore en isolant une partie du réseau qui interagit très peu
avec le reste de la communauté.
La
pyramide trophique.
On a dit, qu'en
fonction de leur rôle de producteur ou de consommateur, les espèces
ou les groupes d'espèces peuvent être affectés à différents niveaux
trophiques. Graphiquement, on représente ces niveaux sous forme de bandes
horizontales posées les uns sur les autres, et dont la longueur est fonction
de grandeurs telles que la biomasse représentée par tel ou tel niveau,
ou bien la quantité d'énergie qui peut être associée à chacun des
niveaux.
Lors qu'on envisage
l'une ou l'autre de ces caractéristiques, on constate que la biomasse,
le nombre d'organismes ou l'énergie sont moindres dans un niveau donné
par rapport au précédent, si bien
que le graphique, large à la base, se rétrécit à chaque niveau supplémentaire
pour donner à la figure l'aspect d'une pyramide à degrés. On appelle
cette figure une pyramide trophique.
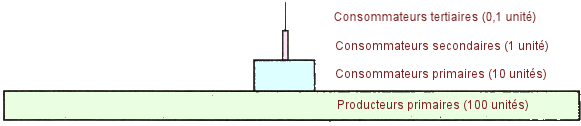
| Pyramide
trophique à quatre niveaux. - Chaque organisme d'une chaîne alimentaire
peut être placé dans une pyramide trophique, où les producteurs sont
situés en bas et les consommateurs tertiaires en haut. La plupart des
pyramides trophiques ont trois niveaux de consommateurs. Au fur et Ă mesure
que l'on monte dans la pyramide, on applique la règle des 10 %, (voir
ci-dessous) qui stipule qu'Ă chaque niveau 90 % de l'Ă©nergie utilisable
est perdue dans l'environnement sous forme de chaleur et que seulement
10 % de l'énergie utilisable est disponible pour le niveau supérieur.
Ci-dessous,
une pyramide trophique imagée et idéalisée. (Source
: M. Castelluci, Science, 2003). |
La
déperdition de l'énergie.
Les chaînes alimentaires
d'un réseau trophique comportent rarement plus de sept maillons (niveaux
trophiques) entre le producteur et n'importe quel prédateur supérieur.
La plupart des chaînes alimentaires sont constituées quatre maillons,
parfois cinq.
•
La principale raison en est l'inefficacité du transfert d'énergie
d'un niveau trophique au suivant (hypothèse énergétique). L'énergie
est perdue sous forme de chaleur (suite à l'activité des organismes)
entre chaque niveau trophique en raison de la deuxième loi de la thermodynamique.
Les chiffres peuvent être différents d'un niveau trophique à un autre
et selon l'écosystème considéré, mais on peut estimer qu'en moyenne
seulement 10 % environ de l'énergie stockée dans la matière organique
de chaque niveau trophique est convertie en substance organique au niveau
trophique suivant. Il s'ensuit qu'après un nombre limité de transferts
trophiques d'énergie, la quantité d'énergie restante dans la chaîne
alimentaire peut ne pas ĂŞtre assez importante pour soutenir des populations
viables à un niveau niveau trophique supérieur.
-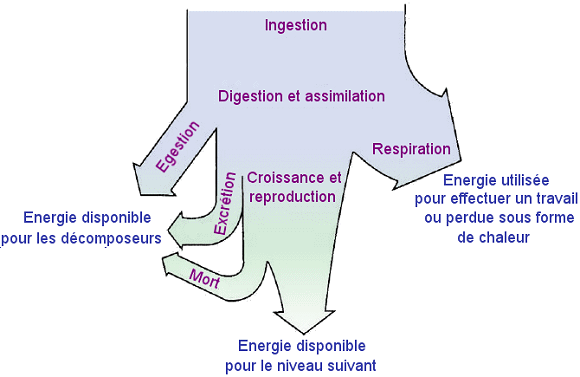 Distribution
de l'Ă©nergie dans chaque niveau trophique.
Distribution
de l'Ă©nergie dans chaque niveau trophique.
• Une autre
raison peut aussi être invoquée : les chaînes alimentaires longues sont
moins stables que les chaînes courtes (hypothèse de stabilité dynamique).
Les fluctuations de population aux niveaux trophiques inférieurs sont
amplifiées aux niveaux supérieurs et peuvent conduire à l'extinction
locale des prédateurs supérieurs. Pour les grands prédateurs, plus la
chaîne alimentaire est longue, et plus la récupération des aléas environnementaux
est lente et difficile.
L'amplification
biologique.
Indépendamment
de la déperdition d'énergie, d'autres phénomènes peuvent s'observer
lors du passage d'un niveau trophique au niveau supérieur. L'un des plus
critiques est l'amplification biologique ou bioamplification,
qui correspond Ă la concentration croissante de substances persistantes
et toxiques dans les tissus des organismes (bioaccumulation) Ă
la suite de l'ingestion d'espèces du niveau trophique précédent, depuis
les producteurs primaires jusqu'aux consommateurs supérieurs. De
nombreuses substances sont susceptible d'une telle accumulation pouvant
atteindre le seuil de toxicité pour les consommateurs des niveaux trophiques
les plus élevés. Parmi les substances susceptibles d'une bioamplification
problĂ©matique, on a identifiĂ© de longue date plusieurs pesticides, Ă
commencer par le DDT(dichlorodiphényltrichloroéthane) et le chlordécone,
ou les BPC (biphényles polychlorés) et les métaux lourds (plomb,
mercure, cadmium, etc.).
Les cycles biogéochimiques.
Les processus trophiques
impliquent un flux d'énergie et de matière. L'énergie finit par être
dissipée sous forme de chaleur et est perdue à jamais pour les processus
métaboliques. Les éléments chimiques absorbés par les organismes vivants
(eau, carbone, azote, phosphore, soufre, etc.) sont appelés, eux, à être
recyclés : ils seront restitués à la composante abiotique de l'écosystème,
soit du vivant même de ces organismes, soit après leur mort sous l'action
des décomposeurs. Il seront alors de nouveau disponibles pour être utilisés
par les organismes vivants. Il suivent donc un cycle, appelé cycle
biogéochimique.
Voici un rapide tour d'horizon des principaux
cycles biogéochimiques. On pourrait encore mentionner les cycles de l'hydrogène,
du potassium, etc.
Le
cycle de l'eau.
La
disponibilité de l'eau influence le rythme de tous les processus à l'oeuvre
dans les écosystèmes terrestres, en contrôlant la production primaire
et la décomposition. De nombreux organismes vivants, comme les plantes,
les animaux et les champignons, dépendent de la très petite quantité
d'eau douce de surface, dont le manque peut avoir des effets considérables.
Les organismes doivent ĂŞtre capables de s'adapter aux fluctuations de
l'approvisionnement en eau.
L'eau liquide est
la phase physique principale de l'utilisation de l'eau, bien que certains
organismes puissent utiliser la vapeur d'eau. Le gel des eaux souterraines
limite la disponibilité de l'eau pour les plantes terrestres. Si l'on
examine les réserves d'eau sur Terre, on constate que 97,5 % sont des
eaux salées non potables. Parmi l'eau restante, 99 % est enfermée dans
le sous-sol sous forme d'eau ou de glace. La quantité d'eau dans l'atmosphère
est minime. Au final, moins de 1% de l'eau douce est facilement accessible
à partir des lacs et des rivières.
Divers processus
régissent le cycle de l'eau. Les principaux sont l'évaporation de l'eau
liquide par l'énergie solaire qui réchauffe les océans et les autres
eaux de surface, la condensation de la vapeur d'eau dans les nuages et
les précipitations, puis l'écoulement des eaux de surface et souterraines
qui renvoie l'eau vers les océans, complétant ainsi le cycle :
•
Evaporation / sublimation : Le cycle de l'eau est alimenté par l'énergie
en provenance du Soleil. Cela entraîne l'évaporation
(eau liquide vapeur d'eau)
des eaux de surface liquides et la sublimation (glace
vapeur d'eau) de l'eau gelée, ce qui libère de grandes quantités de
vapeur d'eau dans l'atmosphère.
• Condensation
/ précipitation : au fil du temps, cette vapeur d'eau se condense en nuages
sous forme de gouttelettes liquides ou gelées et est finalement suivie
de précipitations (pluie ou neige),
qui ramènent l'eau à la surface de la Terre.
• Ecoulement
de l'eau sous la surface : La pluie finit par pénétrer dans le sol, où
elle peut s'Ă©vaporer Ă nouveau si elle est proche de la surface, s'Ă©couler
sous la surface ou être stockée pendant de longues périodes.
• Ruissellement
de surface / fonte des neiges : Le ruissellement
de surface, c'est-Ă -dire l'Ă©coulement d'eau douce provenant de la pluie
ou de la fonte des glaces, est plus facile Ă observer.
• Ecoulement
fluvial : Les eaux de ruissellement peuvent ensuite emprunter des cours
d'eau et des lacs pour atteindre les océans ou s'écouler directement
dans les océans eux-mêmes.
La pluie et le ruissellement
de surface sont les principaux moyens par lesquels les minéraux, notamment
le carbone, l'azote, le phosphore et le soufre, passent de la terre Ă
l'eau.
Le cycle de l'eau
existerait encore en l'absence d'organismes vivants sur la Terre, mais
la transpiration des plantes terrestres qui mobilise un volume d'eau important
lui confère son caractère particulier.
Le
cycle du carbone.
Le carbone est le
deuxième élément le plus abondant dans les organismes vivants. Présent
dans toutes les molécules organiques, il en forme le squelette et
est aussi au coeur des processus par lesquels l'énergie circule à l'intérieur
des écosystèmes. Par ailleurs, lié à l'oxygène, il forme le CO2
atmosphérique, qui est le gaz ayant la plus grande part dans l'effet
de serre (L'atmosphère
de la Terre). Il joue Ă ce titre un rĂ´le pivot dans les Ă©quilibres
à l'intérieur de la biosphère.
La description du
cycle du carbone peut se faire en se plaçant à deux échelles temporelles
différentes, celle, courte, des organismes vivants, puis celle de la biosphère
qui implique de longues durées-:
•
Les organismes photosynthétiques (autotrophes) utilisent le dioxyde de
carbone (CO2) atmosphérique lors de la photosynthèse
et, tout en libérant de l'oxygène, convertissent le carbone en formes
organiques qui sont utilisées par tous les hétérotrophes.
La
respiration aérobie nécessite de l'oxygène provenant de l'atmosphère
ou dissous dans l'eau. Il existe ainsi un échange constant d'oxygène
et de dioxyde de carbone entre les autotrophes (qui ont besoin de carbone)
et les hétérotrophes (qui, en plus, ont besoin d'oxygène).
• Le carbone
utilisé par les organismes vivants se retrouve ensuite dans le sol et
dans les sédiments terrestres (où il forme notamment les grandes réserves
de combustibles fossiles, qui sont les restes décomposés en anaérobie
de plantes et qui mettent des millions d'années à se former). Sur terre,
le carbone est stocké dans le sol à la suite de la décomposition d'organismes
vivants par des décomposeurs ou de l'altération des roches et des minéraux
terrestres. Ce carbone peut être aussi lessivé dans les réservoirs d'eau
par le ruissellement de surface.
+ L'Ă©change
de carbone entre l'atmosphère et les réservoirs d'eau influence la quantité
de carbone trouvée en chaque lieu, et chacun affecte l'autre réciproquement.
Le dioxyde de carbone de l'atmosphère se dissout dans l'eau et se combine
avec les molécules d'eau pour former de l'acide carbonique, puis il s'ionise
en ions carbonate et bicarbonate.
La partie du carbone
fixée dans les roches sédimentaires
carbonées telles que le calcaire du fond de l'océan est ensuite entraînée
dans les profondeurs de la Terre par la subduction des plaques
tectoniques. Le volcanisme sera, plus tard, responsable de son retour
dans l'atmosphère, bouclant ainsi le cycle.
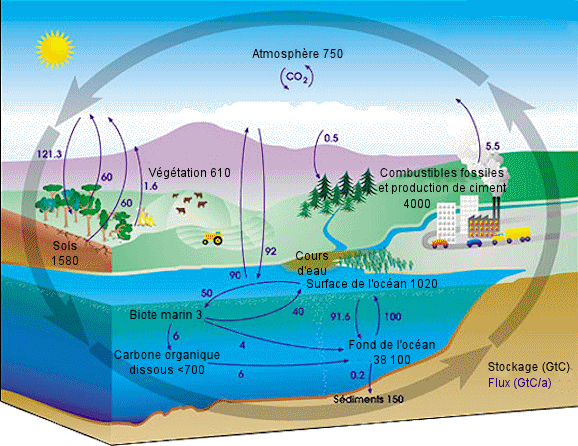
| Le
cycle du carbone. - Chaque année, des dizaines de milliards de tonnes
de carbone circulent entre l'atmosphère, l'hydrosphère et la lithosphère.
Les activités humaines ajoutent environ 5,5 milliards de tonnes par an
de dioxyde de carbone dans l'atmosphère. On a mentionné en noir les quantités
totales de carbone stocké en gigatonnes de carbone (GtC), et, en violet,
les flux annuels de carbone en gigatonnes de carbone par an (GtC/a).
Source : Nasa Earth Observatory. |
Le
cycle de l'oxygène.
L'oxygène est surtout
contenu dans les minéraux de silicate et d'oxydes
de la lithosphère et du manteau terrestre
(99,5 %). Sa présence, comparativement bien moindre, dans l'atmosphère
terrestre est en lien direct avec les cycles de l'eau et du carbone.
Un petite partie
de cet oxygène atmosphérique est issue de la dissociation par le rayonnement
ultraviolet du Soleil des molécules d'eau et de CO2,
mais l'essentiel provient de la photosynthèse,
qui est une photolyse de l'eau effectuée par les plantes et le phytoplancton,
et aussi dans une très large mesure par une cyanobactérie marine (le
Prochlorococcus, qui est le plus petit et le plus abondant organisme photosynthétique
de la planète), à l'origine de la moitié de l'oxygène présent dant
l'atmosphère actuelle de la Terre.
L'oxygène est fortement
réactif, il se lie rapidement à d'autres éléments (oxydation). Il est
ainsi retiré en permanence à l'atmosphère par des réactions abiotiques
(fixation de l'oxygène libre par diverses réactions aux roches de surface)
et biotiques, comme la respiration et la décomposition qui aboutissent
à la libération dans l'atmosphère de dioxyde de carbone. Si l'atmosphère
parvient à maintenir un niveau de 21% d'oxygène, elle le doit seulement
à l'activité photosynthétique des organismes vivants qui y injectent
aussi en permanence ce gaz.
Le
cycle de l'azote.
L'azote est un composant
des acides aminés, des protéines et des acides
nucléiques, et est un élément nutritif essentiel et particulièrement
important pour l'étude de la dynamique des écosystèmes, puisque de nombreux
processus, tels que la production primaire et la décomposition, sont limités
par l'apport d'azote disponible.
Le principal réservoir
d'azote est l'atmosphère, composée à 78 % d'azote gazeux (N2).
Les autres réservoirs sont dans le sol, la biomasse, les sédiments au
fond des lacs, des cours d'eau et des océans, ou encore, sous forme dissoute,
les eaux de surface et souterraines.
Les producteurs primaires,
tels que les plantes et le phytoplancton, qui ordinairement font entrer
les nutriments dans la chaîne trophique ne sont pas capables d'incorporer
l'azote de l'atmosphère. La principale voie d'entrée de l'azote dans
les écosystèmes est donc plutôt la fixation du N2
par des bactéries (bactéries nitrifiantes),
telles, par exemple, les Rhizobium, qui vivent en symbiose dans les nodules
racinaires des légumineuses, ou les Azobacter,
qui sont des bactéries libres. Ces bactéries lient l'azote à l'oxygène
pour former l'ion nitrate (NO3-), qui associé
Ă divers anions (ions positifs), forme les nitrates proprement dits. Ces
composés azotés organique peuvent alors être absorbés aisément par
les plantes.
Dans la suite du
cycle, l'azote sera réintroduit dans l'atmosphère par d'autres bactéries
ou par des champignons sous forme d'azote gazeux. Ce processus se déroule
en trois étapes dans les systèmes terrestres : ammonification, nitrification
et dénitrification.
•
L'ammonification convertit les déchets azotés d'animaux vivants ou de
restes d'animaux morts en ammonium (NH4+)
par certaines bactéries et champignons.
• La nitrification
est la conversion de NH4+ en NO3-.
L'ammonium est d'abord converti en nitrites (NO2-)
par des bactéries nitrifiantes, telles que Nitrosomonas. Les nitrites
sont ensuite convertis en nitrates (NO3-)
par des organismes similaires.
• La dénitrification
intervient dans des conditions anaérobies-:
les bactéries nitrifiantes, telles que Pseudomonas et Clostridium, utilisent
du NO3- au lieu de l'O2
pour leur métabolisme et libèrent du N2 (azote gazeux)
qui peut réintégrer l'atmosphère.
Le cycle de l'azote
océanique se déroule de façon très similaire par l'intermédiare aussi
de bactéries. Une partie de cet azote tombe au fond de l'océan sous forme
de sédiments que les mouvements géologiques finissent par faire revenir
en surface, où l'azote présent peut éventuellement être réinjecté
dans les écosystèmes terrestres.
Cycle
du phosphore.
Le phosphore est
un constituant essentiel des acides nucléiques,
des phospholipides, de l'ATP et d'autres molécules
de stockage d'Ă©nergie; sous la forme de phosphate de calcium, il constitue
les composants de soutien des os. NĂ©cessaire Ă la
croissance des Poissons, le phosphore est souvent le nutriment limitant
dans les écosystèmes aquatiques.
Aucun gaz atmosphérique
ne contient du phosphore (cet élément n'est éventuellement présent
dans l'atmosphère que sous forme de poussière minérale, d'aérosols
ou de cendres volcaniques). En renvanche, le phosphore se rencontre en
grandes quantités dans le sol ou dissous dans les océans, ainsi
que dans les organismes vivants. Mais ce sont les roches sédimentaires
d'origine marine qui constituent le principal réservoir de phosphore :
il y est issu des corps des organismes océaniques et de leurs excrétions.
La seule forme de
phosphore matière inorganique d'importance biologique est l'ion phosphate
(PO4+), que les plantes absorbent et utilisent
pour synthétiser des composés organique ensuite distribués dans tout
le réseau trophique. Le phosphate retourne au sol ou à l'eau par la décomposition
de la biomasse ou l'excrétion par les consommateurs. L'humus et les particules
de sol lient alors les phosphates, les rendant de nouveau disponibles pour
les producteurs primaires.
Le cycle du phosphore
a ainsi tendance à être largement localisé dans les écosystèmes. Mais
le parcours du phosphore peut aussi ĂŞtre beaucoup plus long. Une partie
des phosphates contenus dans les roches est lessivée par les eaux souterraines
et de surface et atteint finalement les océans. Le retour du phosphate
de l'océan vers la terre et à travers le sol est ensuite extrêmement
lent : l'ion phosphate a un temps de séjour
océanique moyen compris entre 20.000
et 100.000 ans.
Cycle
du soufre.
En tant que composant
de la cystéine (un acide aminé), le soufre est impliqué, au sein des
protéines,
dans la formation de liaisons disulfure, qui contribuent à déterminer
leur repliement tridimensionnel, et donc leurs fonctions. Par ailleurs,
certains écosystèmes reposent sur des chimioautotrophes utilisant le
soufre (sous forme de sulfates) comme source d'Ă©nergie biologique.
Présent dans l'atmosphère
sous forme de dioxyde de soufre (SO2), le soufre y
pénètre de trois manières : par la décomposition de molécules organiques,
par l'activité volcanique et les évents géothermiques, et par la combustion
de combustibles fossiles par les humains. Il se dépose ensuite sur la
terre aussi de trois manières principales :
•
Lorsque la pluie tombe, le dioxyde de soufre atmosphérique se trouve dissous
sous forme d'acide sulfureux (H2SO3).
• Le soufre
peut également tomber directement de l'atmosphère.
• De plus,
l'altération des roches contenant du soufre libère du soufre dans le
sol. Ces roches proviennent de sédiments océaniques qui sont déplacés
vers la terre par le soulèvement géologique de ces sédiments.
Les écosystèmes terrestres
utilisent ces sulfates du sol sous forme liquide. L'ion sulfate (SO4--)
est réduit par les plantes et les autres producteurs primaires qui l'incorporent
à leurs protéines sous forme solide. Ensuite, les consommateurs des ces
plantes l'intègrent à leurs protéines. Lors de la mort et de la
décomposition de ces organismes, le soufre réduit des protéines est
oxydé par les bactéries et de nouveau assimilable par les plantes, ou
bien il est rejeté dans l'atmosphère sous forme de sulfure d'hydrogène
(H2S).
Le soufre pénètre
dans l'océan par le ruissellement de la terre, par les retombées atmosphériques
et par les évents géothermiques sous-marins.
Les
effets de l'activité humaine sur les cycles biogéochimiques.
En raison des capacités
technologiques des humains et des systèmes de représentation dans lesquels
ils inscrivent leurs existences individuelles, qui induisent chez eux des
besoins qui vont bien au-delĂ de la dimension simplement biologique, les
humains ont, sur l'écosphère, un impact collectif bien plus grand et
bien plus varié que celui des autres animaux. On se contentera ici d'évoquer
l'impact des activités humaines sur les cycle biogéochimiques.
Afin d'accroître
la disponibilité de l'eau à leur bénéfice, les humains modifient
le cycle de l'eau de diverses manières (collecte d'eaux souterraines,
barrages sur les cours d'eau, irrigation, recours au dessalement pour obtenir
de l'eau potable à partir de l'océan, etc).
-

| Le
fond asséché de la mer d'Aral en Ouzbékistan. - Autrefois alimenté
en eau par deux fleuves importants, l'Amou-Daria et le Syr-Daria, ce lac
d'Asie centrale est aujourd'hui en grande partie asséché. Cette catastrophe
écologique majeure est due au du détournement des deux fleuves, dans
la seconde moitié du XXe siècle, pour servir à l'irrigation de grandes
plantations de coton. Photo : Anton Ruiter; licence
; Creative Commons. |
Les humains interviennent
de manière plus cruciale encore dans le cycle du carbone, en partie du
fait de l'élevage de bovins, source méthane (CH4),
mais principalement depuis le début de l'ère industrielle, par l'injection
dans l'atmosphère de quantités massives de dioxyde de carbone issu de
la combustion des combustibles fossiles. La présence croissante dans l'atmosphère
de CH4 et CO2, qui sont des gaz
à effet de serre, affecte profondément le climat et bouleverse tous les
écosystèmes. Les océans, qui absorbent environ un quart du CO2
atmosphérique, contribuent à limiter jusqu'à un certain point cet effet
de serre, mais en contrepartie ils s'acidifient, ce qui met en péril les
écosystèmes marins. L'acidification des océans a augmenté de 26.%
depuis le début de l'ère industrielle et ne fait que s'accélérer.
La combustion de
combustibles fossiles libère aussi des oxydes d'azote (oxyde nitreux,
N2O, en particulier) qui sont aussi des gaz Ă effet
de serre. Mais les rejets de composés azotés par les activités humaines
sont surtout reliés à l'agriculture. Pour faciliter la croissance et
la productivité des plantes cultivées, on leur apporte souvent des nutriments
en abondance, sous forme d'engrais. Ces engrais contiennent notamment de
l'azote (nitrates), du phosphore (phosphates) et du potassium, et les cycles
de ces éléments sont eux aussi fortement affectés. Les excès de ces
éléments pénètrent dans les sols ou sont véhiculés par le ruisselement
de surface jusqu'aux rivières, aux lacs et aux océans. Ils provoquent
alors une croissance excessive de micro-organismes (processus d'eutrophisation),
qui épuisent l'oxygène dissous, ce qui entraîne la mort de la faune
de nombreux écosystèmes. Ce processus est responsable des zones mortes
(zones incapables de supporter des organismes vivants) dans les lacs et
Ă l'embouchure de nombreux grands fleuves. On estime que ces zones mortes,
dont certaines existent aussi en plein océan (peut-être, dans ce cas,
pour des raisons liées au réchauffement climatique) représentent aujourd'hui
dans le monde une superficie de l'ordre de 250 000 km², soit plus que
celle du Royaume-Uni et près de la moitié
de la superficie de la France métropolitaine
Enfin, le cycle du
soufre est lui aussi fortement impactĂ© par les activitĂ©s humaines, Ă
commencer à par le recours aux combustibles fossiles (pétrole, gaz naturel,
charbon) qui libère dans l'atmosphère du dioxyde de soufre (SO2)
et de sulfure d'hydrogène (H2S). L'eau de pluie
tombant sur le sol à travers le sulfure d'hydrogène et le dioxyde de
soufre les transforme en acide sulfureux (H2SO3).
Il en résulte des pluies acides toxiques qui abaissent le pH des lacs
(ce qui tue une grande partie de la faune résidente); les pluies acides
impactent également directement les humains en affectant leur santé et
en dégradant chimiquement les bâtiments. (Ajoutons que les plus acides
peuvent avoir d'autres origines, notamment du fait de rejets de composés
azotés, tels que l'acide nitrique, HNO3).
Les communautés biologiques
On nomme communauté
(= biocénose) l'ensemble des espèces qui cohabitent et interagissent
au sein d'un même écosystème. Les communautés sont des entités complexes
définies par leur structure (les types et le nombre d'espèces) et leur
dynamique (comment elles changent au fil du temps).
Les interactions
biologiques.
Dans une communauté, les interactions
entre individus de deux espèces (individus hétérospécifiques) ou d'une
même espèce (individus conspécifiques) peuvent prendre plusieurs formes.
On parle de neutralisme dans le cas oĂą aucune des deux ne subit
ni de préjudice ni de bénéfice du fait de l'interaction. L'amensalisme
correspond à la situation dans laquelle la relation cause un préjudice
Ă l'une, mais a un effet neutre pour l'autre. On parle enfin d'antagonisme
quand l'interaction bĂ©nĂ©ficie Ă une espèce et est prĂ©judiciable Ă
l'autre. On parle de facilitation quand au moins l'un
des espèces tire bĂ©nĂ©fice de l'interaction sans porter prĂ©judice Ă
l'autre (mutualisme, commensalisme).
Une interaction entre
deux organismes est dite obligatoire lorsque au moins l'un des deux
organismes ne pourrrait vivre l'un sans interaction avec l'autre. Elle
est dit facultative, quand l'un ces organismes sont capables
de survivre sans cette interaction. Certaines interactions sont de courte
durée (par exemple la prédation, qui est instantanée, la pollinisation;
la zoocorie), d'autres de longue durée, comme le mutualisme, le parasitisme,
la compétition, le neutralisme, etc.
Les principaux
types d'interactions biologiques
| Espèce |
Antagonisme |
Mutualisme |
Amensalisme |
Commensalisme |
Neutralisme |
| I |
+ |
+ |
= |
= |
= |
| II |
- |
+ |
- |
+ |
= |
| Le
signe + signifie que l'interaction est bénéfique pour les individus de
l'espèce concernée; le signe - signifie que l'interaction a des effets
néfastes sur ces individus; le signe = signifie que l'impact de l'interaction
n'est ni positif ni négatif. |
Antagonisme
(+ -).
Les interactions
antagonistes sont celles dans lesquelle l'interaction favorise un organisme
ou une espèce au détriment d'un autre organisme ou d'une autre espèce.
Le comportement associé à ce type d'interaction est appelé exploitation(d'une
espèce par l'autre). Les espèces préjudiciées ont souvent développé
au cours de leur évolution diverses stratégies de défense qui peuvent
être mécaniques, chimiques, comportementales, etc., tels sont le mimétisme,
les piquants, la toxicité, les carapaces, la fuite ou, au contraire, l'immobilité,
etc.
•
La
prédation est l'interaction entre animaux,
dans laquelle un individu d'une espèce (le prédateur) se nourrit, après
capture, d'un individu d'une autre espèce (la proie), qui ne survit pas
Ă l'interaction.
• L'herbivorĂe
(ou phytophagie) est l'interaction dans laquelle un animal ne nourrit
d'un végétal. Elle peut se décliner de diverses
façons, selon les végétaux ou les parties de végétal consommés
: herbivorie proprement dite (herbe); phyllophagie ou folivorie (feuilles);
frugivorie (fruits); granivorie (graines).
Chaque terme a son pendant dans la qualification de l'animal selon son
mode d'alimentation (herbivore, phyllophage; frugivore, granivore, etc.).
• Le parasitisme
est une interaction dans laquelle l'organisme d'une espèce (le parasite)
vit à la surface (ectoparasitisme), à l'intérieur (endoparasitisme)
ou en partie en surface et en partie à l'intérieur (mésoparasitisme)
d'un organisme d'une autre espèce (l'hôte) pour en tirer un bénéfice,
généralement à son détriment (parfois pouvant même entraîner la mort
de l'hôte). Le bénéfice tiré peut être divers. Le parasite peut se
nourrir grâce à son hôte, ou avoir besoin de ce dernier pour se reproduire,
comme dans le cas des virus. Cette interaction
peut être facultative pour certaines espèces de parasites, mais elle
obligatoire pour d'autres.
Le
coopération et le mutualisme (+ +).
La coopération
est une interaction ayant des effets bénéfiques réciproques pour des
organismes d'espèces différentes ou appartenant à la même espèce.
Dans le cas de coopération entre deux espèces distinctes, on parle de
mutualisme.
Le mutualisme est une interaction à long terme. La symbiose et le protocoopération
sont deux types de mutualisme :
•
La
symbiose est une forme de mutualisme entre deux espèces dans laquelle
tous ou quelques-un des individus concernés sont bénéficiaires. C'est
une interaction obligatoire pour l'une des espèce concernées, à au moins
un stade de sa vie. La symbiose est dite unilattérale quand le
caractère obligatoire ne concerne que l'une des espèces; elle est bilatérale
quand l'interaction est obligatoire pour les deux. On appelle holobionte
le tout constitué par des organismes vivant en symbiose.
• La protocoopération
est une interaction rendant sont mutuellement bénéficiaires les individus
ou les populations qu'elle concerne, sans que cette relation soit essentielle
pour l'une ou l'autre. Deux exemples : La pollinisation d'une plante
phanérogame par un insecte, la mycorhyse entre entre
un champignon et les racines d'une plante.
Amensalisme
(= -).
L'amensalime correspond
à un type d'interaction de longue durée entre organismes, qui n'a ni
bénéfice ni préjudice pour l'un, mais qui est préjudiciable à l'autre.
On distingue principalement deux formes d'amensalisme, l'allélopathie
négative et la
compétition :
•
L'allélopathie
négative est une interaction biochimique entre deux organismes, dans
laquelle un organisme est néfaste à un autre en produisant et diffusant
certains composés chimiques.
•
La
compétition est une interaction dans laquelle la présence d'un organisme
dans un une zone réduit ou prive complètement un autre organisme de ressources
vitales (nourriture, lumière), qui sera dès lors exclu de cet habitat.
De ce point de vue la compétition, qui peut aussi être préjudiciable
aux deux individus ou espèces en précence (- -) peut également être
rangée parmi les antagonismes.
Le
principe d'exclusion compétitive. - Ce principe stipule que, dans un habitat
donné, deux espèces ne peuvent pas occuper la même niche écologique
: différentes espèces ne peuvent pas coexister dans une communauté si
elles sont en concurrence pour les mĂŞmes ressources.

| Un
Renard et un Blaireau occupés à se nourrir de prunes tombées de l'arbre.
- Ces deux espèces, classées parmi les Carnivores, sont omnivores en
pratique. Elles peuvent partager le même habitat, où elles sont en compétition
pour l'accès à la nourriture (occasionnellement de façon très flagrante),
mais, malgré des chevauchements possibles pour lesquels il existe une
compétition, elles n'occupent pas exactement la même niche écologique
(partitionnement de niche), ce qui assure leur coexistence durable. Photo
: © Serge Jodra, 2022. |
Le
commensalisme (= +).
Contrairement Ă
l'amensalisme, le commensalisme (au sens large) désigne une des interactions
entre organismes qui, si elles n'ont ni bénéfice ni préjudice pour l'un
(appelé l'hôte), ont un effet bénéfique sur l'autre (le commensal,
l'inquilin, le phoronte, l'Ă©pibionte, etc).
•
Commensalisme
proprement dit. - Au sens propre, le commensalisme désigne
les interactions dans lesquelles une espèce prĂ©lève sa nourriture Ă
partir d'une autre. L'Ă©tymologie (le mot vient du latin cum = avec
et mensa = table), rappelle que le commensal est celui qui mange
"Ă la mĂŞme table" que son hĂ´te.
On peut y rattacher
d'autres interactions telles que :
•
L'inquilinisme,
qui concerne les situations dans lequelles une espèce est hébergée par
un hôte auquel elle ne nuit ni de bénéficie. Par exemple, quand un oiseau
utilise le nid d'un autre après que celui-ci l'ait
abandonné.
• L'allélopathie
positive, qui est une interaction biochimique entre deux organismes,
dans laquelle un organisme bénéficie à un autre en produisant certains
composés chimiques. L'allélopathie, qu'elle soit positive ou négative,
concerne essentiellement les plantes et les micro-organismes.
• L'épibiose,
qui correspond à l'interaction entre une espèce et une autre espèce
qui lui sert de support (cas des mousses , des
lichens,
des orchidées). Contrairement au parasite,
l'épibionte ne prélève rien au détriment de son hôte.
• La phorésie,
qui renvoie à des situations où une espèce se sert d'une autre comme
moyen de transport. Exemple, les rémoras qui vivent accrochés aux requins.
• La zoocorie
est une interaction qui peut être rapprochée de la précédente, et par
laquelle un animal transporte les graines des plantes et contribue ainsi
à la dissémination des semences. Dans
l'exozoocorie, les graines adhèrent à la peu de l'animal; dans
l'endozoocorie, les graines sont absorbées par l'animal, puis rejetées
dans ses excréments.
• La thanatochrèse
est l'utilisation de restes, d'excréments, de squelettes ou de cadavres
d'autres espèces afin de se protéger ou de les utiliser comme outils.
Par exemple, les pagures (bernard-l'hermite), qui utilisent des coquilles
de gastéropodes pour protéger leur abdomen.
La biodiversité.
La biodiversité
ou diversité biologique définit la complexité biologique d'une communauté.
Cette diversité a deux composantes :
•
La
richesse en espèces (richesse spécifique), qui est le nombre d'espèces
différentes vivant dans dans une zone particulière, un habitat,
un biome ou même la biosphère dans son entier. La plus grande richesse
en espèces s'observe dans les écosystèmes proches de l'équateur
(températures plus chaudes, grandes quantités de précipitations, faible
saisonnalité); à l'inverse, la plus faible richesse en espèces se rencontre
aux très hautes latitudes (températures très
basses, faibles précipitations).
• L'abondance
relative des espèces (régularité spécifique), dans une zone particulière,
qui est le nombre d'individus d'une espèce par rapport au nombre total
d'individus de toutes les espèces dans cette zone. Si l'on a deux communautés
de même richesse en espèces, on considère que la biodiversité est plus
grande dans celle où l'abondance relative de chaque espèce est la plus
grande; il y a moins de biodiversité lorsque certaines espèces sont très
représentées et d'autres très peu.
La richesses et l'abondance
relative des différentes espèces qui composent une communauté sont importantes
pour l'intégrité et souvent même la survie de la communauté (on a dit
plus haut que plus un système est complexe et plus il dispose de leviers
sur lesquels il peut agir pour se réguler). Les communautés diversifiées
protègent les écosystèmes en fournissant des services tels que l'ombre,
la dégradation des déchets et la production d'oxygène.
Le succès et la
survie d'une espèce dépendent aussi de sa diversité génétique, c'est-à -dire
de la variété et de la fréquence relative des différents allèles
dans son patrimoine génétique. La diversité génétique peut être essentielle
pour qu'une espèce puisse s'adapter à des environnements changeants.
Espèces
au rĂ´le particulier.
Au sein d'une communauté,
certaines espèces exercent un fort contrôle sur la structure de la communauté,
en particulier sur la composition des espèces présentes, leur abondance
relative et sur la diversité en général. Soit en raison de leur abondance,
soit parce qu'elles jouent un rĂ´le fondamental dans la dynamique de cette
communauté, l'impact de ces espèces peut s'exercer par des interactions
trophiques ou par leurs influences sur le milieu physique. On distingue
ainsi :
•
Les
espèces fondatrices. - Les espèces fondatrices sont considérées
comme le socle sur lequel se constitue une communauté; ce sont celles
qui ont la plus grande influence sur sa structure globale. Elles possèdent
souvent l'abondance relative la plus élevée. Ce sont généralement les
producteurs primaires : des organismes qui apportent la majeure partie
de l'énergie à la communauté. Elles n'exercent leur influence en provoquant
des changements physiques dans l'environnement (soit par leur comportement,
soit par leur importante biomasse) pour produire et maintenir des habitats
qui profitent aux autres organismes qui les utilisent. En modifiant la
structure ou la dynamique de l'environnement, les espèces fondatrices
agissent comme des facilitateurs, avec des effets positifs sur la survie
et la reproduction de certaines autres espèces de la communauté.
•
L'espèce
dominante. - On nomme espèce dominante d'une communauté celle qui
y exerce le plus fort contrôle sur la présence et la distribution d'autres
espèces. C'est ordinairement la plus abondante ou celle qui représente
la plus importante biomasse. L'espèces fondatrice peut ainsi parfois être
l'espèce dominante. Le rôle dominant d'une espèce peut lui venir de
ce qu'elle est plus compétitive dans l'exploitation de ressources limitées
telles que l'eau ou les nutriments. Elle peut ĂŞtre dominante aussi parce
qu'elle est plus efficace pour éviter la prédation ou les conséquences
des maladies.
• Les
espèces clés. - Une espèce clé est une espèce dont la présence
est essentielle au maintien de la biodiversité au sein d'un écosystème
et au maintien de la structure d'une communauté écologique. Contrairement
aux espèces dominantes, les espèces clés n'ont pas besoin d'être abondantes
dans une communauté. Elles exercent un fort contrôle sur la structure
de la communauté d'abord parce qu'elles y jouet un rôle ou y occupent
une niche Ă©cologique fondamentale.
Les successions Ă©cologiques.
Tous les espaces
qui offrent des conditions de vie propices Ă des organismes vivants sont
occupés. Des communautés s'y sont constituées qui y ont acquis une structure
stable. Mais il peut arriver que la composition et la structure communautaire
soient notablement altérées, surtout quand toute la végétation existante
dans la zone est démantelée. Cela peut survenir à la suite de perturbations
environnementales telles que des éruptions volcaniques, des séismes,
des tempêtes, des incendies, la régression de glaciers, l'assèchement
de zones humides, etc. Les effets du changement climatique et l'intervention
humaine jouent ici un rĂ´le majeur. L'Ă©quilibre est rompu parfois temporairement,
parfois définitivement. Dans ce cas, la zone vidée totalement ou partiellement
des espèces qui y vivaient peut être colonisée par de nouvelles espèces,
qui d'ailleurs pourront être progressivement remplacées par d'autres
qui, à leur tour, seront possiblement remplacées par d'autres encore,
jusqu'à ce que s'établisse à nouveau un écosystème avec une communauté
à l'équilibre. Ce processus d'apparition et de disparition séquentielles
au fil du temps d'espèces dans une communauté est appelé succession
écologique. L'état final d'une succession est appelé son climax.
Les successions peuvent
revêtir des caractères différents, selon lien qui existe entre la première
espèces installée et la denière. On peut ici distinguer trois modalités
principales :
•
Les espèces précoces facilitent l'apparition des espèces plus tardives,
leur rendant l'environnement plus favorable (par exemple, en augmentant
la fertilité du sol).
• Les premières
espèces inhibent l'implantation des espèces ultérieures, mais pas suffisamment
pour l'empĂŞcher.
• Les premières
espèces sont totalement indépendantes des suivantes : elles n'aident
ni n'entravent leur colonisation.
Succession
primaire.
Dans la succession
primaire, les terres nouvellement exposées ou nouvellement formées sont
vides de toute espèce vivante et sont colonisées entièrement pas de
nouvelles espèces. Les premiers organismes à s'installer sont les procaryotes
autotrophes.Des organismes photosynthétiques macroscopiques (lichens et
mousses) se développent ensuite à partir de spores
apportés par le vent. Le sol se transforme progressivement,
son pH évolue, à mesure que les roches se détériorent sous l'effet
des intempéries, et que la matière organique s'accumule à partir des
restes en décomposition. Il devient ainsi propice à des installations
nouvelles. Des herbes commencent Ă pousser et recouvrent les mousses et
les lichens. Ces toutes premières espèces, qui contribuent à enrichir
de plus en plus le sol, sont connues sous le nom d'espèces pionnières.
D'autres espèces moins résistantes poussent et remplacent éventuellement
les espèces pionnières : ce peut être des buissons et des arbustes,
dont les graines ont été été apportées par les animaux qui commencent
à occuper ce nouvel espace. Certaines de ce ses nouvelles espèces, apportent
peut-être aussi leurs bactéries symbiotiques, qui fixeront l'azote atmosphérique,
ce qui enrichira encore davantage le sol. Les plantes appelées à devenir
la végétation prédominante du lieu s'installent et se répandent enfin.
Avant que s'Ă©tablisse une structure communautaire stationnaire, il faudra
peut-être encore des siècles, voire plusieurs millénaires.

| L'île
de Surtsey, au large de la côte méridionale de l'Islande.
- Cette île (près d'un kilomètre carré et demi) s'est formée entre
1963 et 1967, Ă la suite d'une Ă©ruption volcanique. Sa dynamique communautaire
( = évolution de la structure et de la composition de la communauté qui
s'y est constituée) est étudiée depuis cette époque. Les premiers insectes
volants, apportés par le vent, y ont été observés dès 1964. Ils ont
fourni une première nourriture à des oiseaux de passage. La suite des
événements météorologiques a eu pour effet de décomposer suffisamment
le substrat pour permettre l'Ă©tablissement de certaines plantes vasculaires
(les premières ont été observées dès 1965) et de lichens nécessitant
peu de sol. Ces espèces pionnières (une vingtaine à Surtsey, dont seulement
la moitié ont persisté) ont aidé à épaissir une couche de matière
organique en décomposition, contribuant ainsi à la formation du sol.
Aujourd'hui, environ 70 espèces de plantes poussent sur l'île. Des mouettes
y ont établi une colonie sur l'île en 1984. Une douzaine d'espèces d'oiseaux
y sont désormais répertoriées. Photo : Harvey Barrison,
2016; licence Creative commons. |
Succession
secondaire.
Dans la succession
secondaire, un écosystème est perturbé gravement, mais des vestiges
de la communauté précédente subsistent. Une telle succession peut aussi
se produire lorsqu'une communauté existante a été supprimée par une
perturbation qui a cependant laissé le sol intact. La zone peut retourner
à un état similaire à son état d'origine. Dans le cas d'une zone forestière
défrichée pour l'agriculture puis abandonnée, des plantes herbacées,
dont les graines ont été apportées par le vent ou les animaux, recolonisent
la zone. Ces plantes sont souvent remplacées par des broussailles
ligneuses, qui elles-mĂŞmes laisseront la place Ă des forĂŞts d'arbres.
Dans le cas, d'un incendie de forêt, la majeure partie de la végétation
disparaît et les animaux incapables de fuir sont tués. Leurs nutriments,
cependant, sont restitués au sol sous forme de cendres. De nouveaux individus
pourront ainsi s'installer dans de bonnes conditions. Avant l'incendie,
la végétation était dominée par de grands arbres ayant accès à la
principale ressource énergétique végétale : la lumière du soleil.
Leur hauteur leur donnait accès à la lumière du soleil tout en ombrageant
le sol et d'autres espèces basses. Après l'incendie, cependant, ces arbres
ne sont plus dominants. Ainsi, les premières plantes à repousser sont
généralement des plantes annuelles suivies en quelques années par des
graminées
à croissance et propagation rapides et d'autres espèces pionnières.
Au fil des ans, des espèces, dites espèces intermédiaires, émergeront
: des arbustes, de petits pins, des chênes, etc. En un ou deux siècles,
la forĂŞt atteindra un Ă©tat d'Ă©quilibre stationnaire.
Les populations
Une population correspond
à l'ensemble des individus de la même espèce vivant dans une zone donnée.
Les membres d'une population dépendent des mêmes ressources, sont influencés
par les mĂŞmes facteurs environnementaux, et sont susceptibles d'interagir
et de se reproduire entre eux. Les populations peuvent évoluer grâce
à l'action de la sélection naturelle sur les variations héréditaires
entre les individus. Les populations fluctuent en fonction de facteurs
biotiques et abiotiques.
Caractérisation
d'une population.
Effectif
et densité d'une population.
Dans un habitat
particulier, une population est caractérisée en premier lieu par son
effectif
( = sa taille, le nombre total d'individus qui la composent), et sa densité
( = le nombre d'individus dans une zone ou un volume donnés).
La
variabilité génétique à l'intérieur d'une population et donc le potentiel
d'adaptation de ses membres Ă l'environnement, explique que les populations
de grand effectif sont souvent plus stables que les populations plus petites.
La
facilité d'un individu à trouver un partenaire pour se reproduire peut
dépendre de la densité de la population à laquelle il appartient. A
cause de cela, les populations à faible densité peuvent avoir plus de
difficultés à se maintenir. Par ailleurs, la disponibilité des ressources
explique que les petits organismes ont tendance à être plus densément
répartis que les grands organismes.
Schémas
de dispersion.
A l'intérieur d'une
zone géographique occupée par une population, sa densité moyenne n'est
pas le seul élément à prendre en compte : les densités locales
peuvent en effet être très diverses. Les membres de l'espèce peuvent
vivre tous très proches les uns des autres ou se trouver éloignés, et
lorsqu'ils sont éloignés les uns des autres, ils peuvent se distribuer
de façons différentes.
On caractérise la
relation spatiale qui existe entre les membres d'une population dans un
habitat à un moment donné en en définissant le
schéma de dispersion
(ou schéma de distribution). Sa connaissance donne des indications
sur les interactions sociales entre les individus de la population étudiée
et sur la manière dont cette population interagit avec son environnement.
Des différences environnementales au niveau local, contribuent aux variations
de la densité de population, car certaines zones de l'environnement conviennent
mieux à l'espèce considérée.
.
Lorsque les membres
d'une population sont concentrés en groupes, uniformément répartis
ou dispersés au hasard, on parle respectivement de de dispersion groupée,
uniforme et aléatoire.
•
La
dispersion groupée, qui est le schéma de dispersion le plus souvent
rencontré, correspond à la concentrations des membres de la population
dans des zones très localisées. On l'observe chez les plantes qui laissent
tomber leurs graines directement sur le sol ou chez les animaux grégaires.
L'hétérogénéité
de l'habitat peut aussi conduire Ă une telle dispersion Les plantes
et les champignons se regroupent souvent dans des endroits oĂą le sol et
d'autres facteurs environnementaux favorisent la germination et la croissance.
Dans les espèces animales, le regroupement peut être un facilitateur
pour l'accouplement, il peut aussi faciliter l'action de certains prédateurs
(chasse en meute des loups, par exemple).
•La
dispersion uniforme est le schéma dans lequel les individus d'une
population sont espacés de manière homogène, généralement avec en
arrière-plan, une stratégie d'accès à des ressources locales limitées.
On l'observe chez les plantes qui sécrètent des substances inhibant la
croissance des individus voisins (allélopathie négative). Des interactions
sociales antagoniques chez certaines espèces animales, comme la défense
d'un territoire, peuvent aussi aboutir Ă une telle dispersion.
• La dispersion
aléatoire, relativement peu commune, correspond à une situation où
l'espacement des membres d'une population n'est pas prévisible-:
la position de chaque individu est indépendante de celle des autres. Ce
schéma de dispersion découle de l'absence de fortes attractions
ou répulsions entre les membres de la population ou lorsque les principaux
facteurs physiques ou chimiques pertinents sont relativement homogènes
dans la zone concernée. On l'observe, par exemple, avec des
plantes dont les graines, disséminées par le vent, germent là où elles
tombent dans un environnement favorable.
Paramètres
démographiques.
La taille et la
densité d'une population décrivent une population à un moment donné.
Mais lorsqu'on étudie une population, il est également nécessaire d'en
considérer l'évolution dans le temps. L'effectif et la densité d'une
population sont les résultantes de divers processus qui lui ajoutent (naissances,
immigration) ou soustraient (décès, émigrations) des membres.
Divers paramètres
permettent de rendre compte de cette Ă©volution. On s'en tiendra ici Ă
ne définir que quelques rapports, utiles pour comprendre la dynamique
d'une population.
•
Le
taux de natalité est le rapport du nombre de naissances vivantes de
l'année à l'effectif total de la population pendant l'année. Le rapport
entre les mâles et les femelles ou la structure d'âge (la proportion
de membres de la population dans des tranches d'âge spécifiques), composée
de nombreux individus en âge de se reproduire peut augmenter le taux de
natalité. Une population nombreuse entraîne un taux de natalité plus
élevé parce qu'il y a plus d'individus potentiellement reproducteurs.
•
Le
taux de reproduction est, dans les espèces qui se reproduisent par
voie sexuelle (et donc où les mâles n'interviennent que pour la distribution
des gènes), le rapport de l'effectif de la progéniture femelle à celui
des mères.
• Le taux
de fécondité. - La fécondité est la capacité de reproduction d'un
individu au sein d'une population; elle décrit le nombre de descendants
qui pourraient ĂŞtre produits si un individu avait autant de descendants
que possible, en répétant le cycle de reproduction aussitôt après la
naissance de la progéniture. En pratique, on définit le taux de fécondité,
qui est le rapport du nombre de naissances au cours de la période de référence
chez les femelles d'un âge donné au moment de la naissance au nombre
de femelles du même âge pendant cette même période
• Le taux
de mortalité est le rapport du nombre de décès de l'année à l'effectif
moyen de la population pendant l'année. On peut y rattacher le temps qu'un
membre de ce groupe est susceptible de vivre (espérance de vie). Une grande
taille de population peut entraîner un taux de mortalité plus élevé
en raison des maladies, de l'accumulation de déchets et de la concurrence
dans un milieu aux ressouces limitées. Dans les populations dépendantes
de la densité, les taux de la mortalité augmentent et les taux de natalité
diminuent lorsque la densité augmente. Dans les populations indépendantes
de densité, ces taux ne changent pas lorsque la densité augmente.
Migrations
et métapopulations.
Les migrations
peuvent aussi jouer un rĂ´le important. Elles peuvent affecter la population
entière qui dès lors change simplement d'habitat (par exemple, la migration
d'un troupeau d'herbivores). La migration peut aussi ne concerner qu'un
nombre défini d'individus, ce qui dans ce cas influe sur l'effectif ou
la densité d'une population donnée.
On parle d'Ă©migration
lorsque des individus appartenant Ă une population la quittent pour rejoindre
une zone géographique différente, et d'immigration lorsque des
individus venus d'une zone différente rejoignent la population considérée.
Le solde migratoire (ou accroissement migratoire) est la
différence entre le nombre d'individus entrées sur un territoire (immigrants)
et le nombre de personnes qui en sont sortis (Ă©migrants), au terme
d'un intervalle de temps donné. Pour évaluer la variation de l'effectif
d'une population (= croissance démographique), on l'ajoute à la
différence entre le nombre de naissances (vivantes) dans la population
et le nombre de décès au cours de ce même espace de temps.
Lorsqu'on considère
ensemble la ou les populations d'origine de migrants et la ou les populations
de destination de ces migrants, on a affaire Ă une population plus vaste,
dont les composantes sont géographiquement distinctes, qui prend alors
le nom de métapopulation.
Stratégies
d'histoire de vie.
Chaque espèce peut
se caractériser par l'histoire de vie de ses représentants, autrement
dit, par les caractéristiques propres au mode de reproduction et la survie
des organismes de cette espèce entre leur naissance et leur mort. Les
histoires de vies varient d'une espèce à l'autre parce que chaque espèce,
confrontée aux limites des ressources (à commencer par l'énergie) que
peut lui procurer son environnement, a développé au cours de l'évolution
diverses stratégies lui permettant d'user des ressources disponibles,
et de les gérer en les répartissant entre les besoins de croissance,
d'entretien et de reproduction, avec la meilleure efficacité. Ces stratégies,
appelĂ©es stratĂ©gie d'histoire de vie, peuvent se rapporter Ă
plusieurs schémas basés principalement sur les trois éléments suivants
:
•
L'âge de la première reproduction ou âge de la maturité. C'est le moment
où la reproduction commence. Il existe une relation entre cet âge et
l'engagement dans les soins parentaux. Les individus d'une espèce peuvent
se reproduire tôt dans leur vie pour s'assurer de survivre jusqu'à l'âge
de la reproduction ou se reproduire plus tard dans leur vie pour devenir
plus grands et en bonne santé et être mieux à même de prodiguer des
soins parentaux.
• La fréquence
de reproduction des organismes. Les individus d'une espèce peuvent se
reproduire une fois (semelparité) ou plusieurs fois (itéroparité) au
cours de leur vie. Le facteur déterminant est ici le taux de survie de
la progéniture.
+ La sémelparité
se produit lorsque les individus d'une espèce ne se reproduisent qu'une
seule fois au cours de sa vie, puis meurent. Les espèces sémelpares utilisent
la majeure partie de leur budget de ressources au cours d'un seul événement
reproductif, sacrifiant leur santé au point qu'elles ne survivent pas.
La sémelparité s'observe souvent chez des espèces vivant dans des environnements
très variables ou imprévisibles.
+L'itéroparité
décrit les espèces dont les représentants se reproduisent à plusieurs
reprises au cours de leur vie. Certains animaux ne peuvent s'accoupler
qu'une fois par an, mais survivent Ă de multiples saisons d'accouplement.
L'itéroparité caractérise surtout des espèces vivant dans un environnement
stable
• Le nombre
de descendants produits dans chaque Ă©pisode de reproduction (nombre de
graines produites par la plante, taille de la portée ou de la couvée
chez les animaux). Le nombre de descendants a souvent un lien avec la fréquence
de reproduction : la descendance est nombreuse dans les environnements
incertains, ce qui augmente la probabilité qu'au moins certains individus
survivront. Dans des environnements plus sûrs, quelques petits, disposant
de provisions suffisantes, auront plus de chances de survivre jusqu'Ă
l'âge de la reproduction. Il existe aussi une relation inverse entre
la fécondité et les soins parentaux, qui sont plus importants quand la
progéniture est peu nombreuse.
Les modèles de la
croissance démographique.
Des modèles mathématiques
peuvent être utilisés pour décrire les changements survenant dans une
population ou pour mieux les prédire. Deux modèles simples et idéalisés
sont présentés ici. Le premier montre que les populations disposant de
ressources illimitées connaissent une croissance exponentielle,
avec un taux de croissance accéléré. Dans le second modèle, on pose
l'existence d'une limite au ressources disponibles et dans ce cas, la croissance
de la population, d'abord rapide, se ralentit puis se stabilise Ă un niveau
maximal. Les situations réelles appellent cependant des approches plus
complexes.
Croissance
exponentielle.
Si l'on ne prend
pas en compte les effets des migrations, à chaque génération l'effectif
d'une population varie en fonction de son taux de natalité
n et
de sonb taux mortalité m. Le taux
r de croissance intrinsèque
de cette population s'Ă©crit alors : r = n - m. La valeur de r
peut être positive (augmentation de l'effectif de la population); ou négative
(diminution de l'effectif de la population; croissance démographique négative),
ou nulle (pas de changement dans la taille de la population; croissance
nulle). Dans le cas d'une croissance négative qui se maintient, la population
est appelée à s'éteindre.
Le taux de croissance
d'une population, c'est-Ă -dire la variation de son effectif N en
un temps t donné est égal à son taux croissance intrinsèque
r
multiplié par cet effectif. En temps continu, on peut exprimer cela sous
la forme d'une équation différentielle :
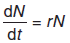
Cette Ă©quation signifie
que le taux de variation de la population N Ă un instant t
(dérivée de N par rapport au temps) est égal au produit du taux
de variation intrinsèque r par l'effectif de la population. Lorsqu'on
l'intègre, on obtient un effectif de la population évoluant en fonction
du temps de façon exponentielle.
Croissance
logistique.
Dans le monde réel,
les ressources ne sont pas illimitées et la croissance exponentielle ne
peut exister (et encore de manière approximative) que tant que les besoins
de la population restent négligeables par rapport aux ressources
disponibles. A partir d'un certain moment, la croissance de la population
est visiblement freinée par l'existence de cette limite : chaque individu
a accès à une quantité décroissante de ressources et il existe une
limite à la quantité d'individus qui peuvent vivrent dans un habitat
donné. Un modèle plus réaliste doit incorporer la notion de capacité
de charge (K), qui est la valeur maximale que peut atteindre une
population dans un environnement donné. Le nombre d'individus pouvant
s'ajouter à une population à un moment donné est K-N. Le rapport
(K-N)/K
correspond alors à la fraction de la capacité de charge disponible pour
une croissance supplémentaire. L'équation précédente doit être modifiée
pour rendre compte de cette contrainte. Elle devient :
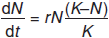
L'intégration de l'équation permet de tracer
une courbe représentative de N en fonction du temps qui prend la forme
d'un sigmoĂŻde, c'est-Ă -dire d'une courbe en forme de S. La droite d'Ă©quation
y = K est son asymptote quand t tend vers l'infini
(Les fonctions).
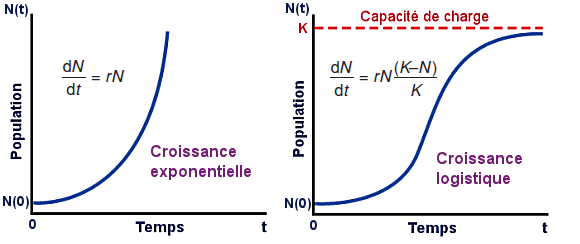
| Croissance
exponentielle et croissance logistique. - L'Ă©quation de la croissance
exponentielle représente la croissance potentielle d'une population dans
un environnement aux ressources illimitées. Ce modèle prédit que plus
une population est grande, plus elle grandit rapidement (rétroaction positive).
L'Ă©quation de la croissance logistique prend en compte l'existence d'une
limite des ressources disponibles. Les premiers temps de la croissance
d'une population soumise Ă telle limitation ressemblent Ă ceux de la
croissance sans limitation de ressources, mais l'existence d'une capacité
de charge se fait vite sentir (rétroaction négative); le taux de croissance
diminue à mesure que la population approche de sa capacité de charge.
La population se stabilise près de cette valeur limite. |
Croissance
logistique et population réelle.
Plus réaliste que
le modèle exponentiel, le modèle logistique de la croissance d'une population
n'en est pas moins une approximation des situations réelles, et il peut
exister bien d'autres manières pour l'effectif d'une population d'évoluer
(cycliques comme dans l'exemple Lièvres-Lynx, ci-dessous; chaotiques;
etc.). Plusieurs facteurs jouent dans l'Ă©volution de l'effectif d'une
population réelle. On a déjà évoqué le rôle des migrations (une émigration
peut intervenir quand les ressources sont plus difficiles d'accès). On
doit aussi tenir compte de la compétition intraspécifique : « tous les
animaux sont égaux, mais certains sont plus égaux que d'autres » (G.
Orwell, 1945) pour accéder aux ressources (cela vaut aussi pour les
populations végétales, bactériennes, etc.). La densification de la population,
qui peut attirer davantage de prédateurs, et l'accumulation de déchets
peuvent aussi être des freins à l'accroissement démographique.
Par ailleurs, le
modèle logistique suppose que les espèces s'adaptent instantanément
Ă la croissance et Ă l'Ă©volution de la population et se rapprochent
en douceur de la capacité de charge. Toutefois, dans la plupart
des populations réelles, il s'écoule un certain temps avant que les facteurs
limitatifs négatifs de la croissance démographique ne deviennent apparents.
Si, par exemple, la disponibilité de la nourriture commence à diminuer
dans une population, la reproduction diminue Ă©galement. Cependant, le
taux de natalité peut ne pas être réduit immédiatement car les femelles
peuvent utiliser leurs réserves d'énergie pour continuer à se reproduire
pendant une courte période. Cela signifie que dans certains cas l'espèce
est en mesure de dépasser sa capacité de charge avant d'atteindre une
densité relativement stable. La capacité de charge peut aussi se modifier
pour des raisons extérieures à la population, ou simplement parce que
les ressources dans lesquelles puise la population ne se renouvellent plus.
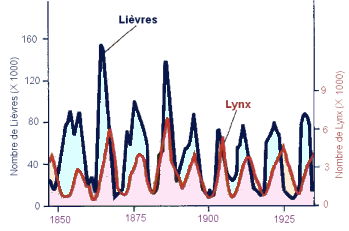
| Cycles
de populations engendrés par une interaction proie-prédateur. - Cet
exemple est tiré de l'étude de l'évolution des populations de Lièvres
américains et de Lynx dans le Nord de l'Ontario
pendant un siècle, à partir du nombre de peaux vendues par les chasseur
de la Compagnie de la baie d'Hudson. Lorsque
le nombre de lièvres augmente, les lynx disposent de plus de nourriture
et leur nombre augmente. Il s'ensuit que le nombre de lièvres diminue
de nouveau. La nourriture venant Ă manquer aux lynx, leur population diminue
à son tour. Moins chassés, les lièvres peuvent de nouveau se multiplier
: le cycle recommence. |
 |
En
librairie. - Collectif, Dictionnaire
de l'Ă©cologie,
Encyclopaedia universalis, 2001; Dieter Heinrich et Manfred Hergt, Atlas
de l'Ă©cologie, Livre de Poche, 1993;
Jean-Paul Deléage, La Biosphère :
Notre terre vivante, Gallimard, 2001;
Robert Barbault, Écologie générale,
Structure et fonctionnement de la biosphère: Structure et fonctionnement
de la biosphère, Dunod, 2021; Jacques
Grinevald, La Biosphère de l'Anthropocène
: Climat et pétrole, la double menace,
Georg Editeur, 2008.
Alain Pavé, Comprendre la biodiversité.
Vrais problèmes et idées fausses: Vrais problèmes et idées fausses,
Seuil, 2019; Christian LevĂŞque, La
biodiversité : avec ou sans l'homme? Réflexions d'un écologue sur la
protection de la nature en France,
QUAE GIE, 2017; Pierre-Henri Gouyon, CĂ©dric Gaucherel, Jean-Louis
Dessalles, Le Fil de la Vie. La face
immatérielle du vivant, Odile Jacob,
2016.
Serge Frontier et Denise Pichod-Viale et al, Écosystèmes,
Structure, Fonctionnement, Évolution,
Dunod, 2008; François Bréchignac et Lisa Cauvin, L'écosystème:
La dimension négligée du vivant,
L'Harmattan, 2022.
Neil Campbell, Jane Reece et al., Biology,
2005; Eugene Odum, Ecology,
1963-1975; P. H. Collin, Dictionary
of environment and ecology, 1992-2004. |
|
|
