| . |
|
||||||
|
|
| . |
|
||||||
| Animaux > Annélides |
| Les
Chétopodes
Oligochètes et Polychètes |
| Les Chétopodes
constituent une superclasse d'Annélides
comprenant des vers qui sont libres, munis de soies paires, implantées
soit dans des cryptes, soit sur des pieds rudimentaires, et possèdent
souvent une tête distincte avec des tentacules ou des cirres.
Les Chétopodes sont surtout des vers marins; quelques-uns vivent dans les eaux douces, d'autres dans la terre humide, comme les lombrics. Certains présentent des phénomènes de reproduction et de métamorphose également singuliers. On range ces vers dans deux classes : Oligochètes, et Polychètes Oligochètes. - Vers sans antennes, ni parapodes, mais pourvus de soies disposées en faisceaux sur chaque segment ou réparties sur un même cercle entourant le segment. Hermaphrodites. Habitent presque tous les eaux douces ou la terre humide.Le corps est divisé en deux parties principales : 1° la tête, appelée aussi lobe céphalique ou prostomium;Extérieurement, cette métamérisation se manifeste le plus souvent par des constrictions qui séparent les divers anneaux et par des bouquets de soies disposés par paires, tantôt dans la paroi du corps, tantôt sur des saillies de cette dernière (parapodes); intérieurement, les métamères sont séparés par des cloisons musculaires transversales (dissépiments), et certains organes sont aussi répétés d'une façon homotypique dans chaque segment (par exemple, les ganglions de la chaîne nerveuse et d'une façon moins complète, les dentonèphres ou organes segmentaires, les glandes génitales, etc.). - 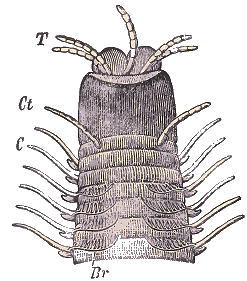
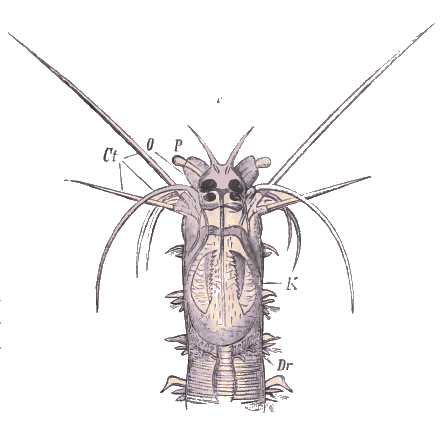
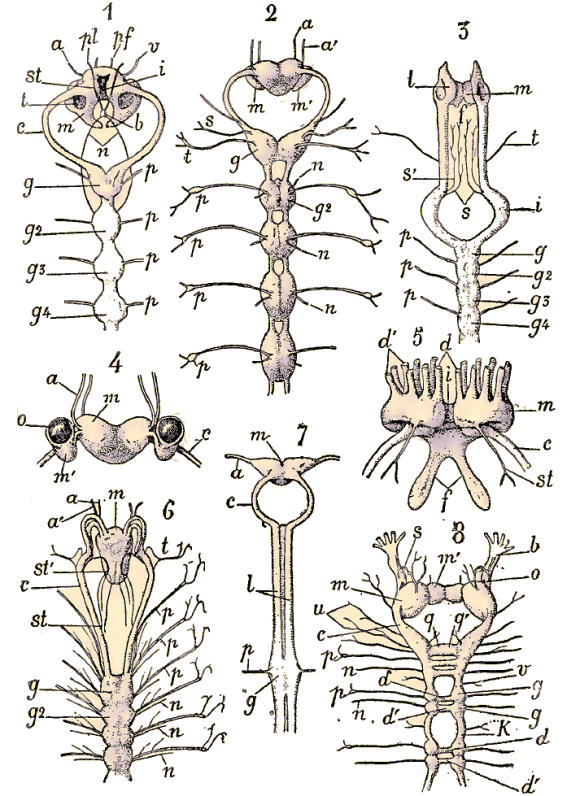
Tête et segments antérieurs d'une Eunice, vue de dos. - T. Antennes; Ct. Cirres tentaculaires; C. Cirres de parapodes; Br. Branchies.
Comme dans toutes les grandes divisions du règne animal, le développement des Chétopodes est tantôt dilaté, tantôt condensé. La forme embryonnaire normale (palingénétique) est la trochosphère ou trochophora. La connaissance de cette forme larvaire est indispensable pour comprendre l'organisation de l'adulte. La trochosphere des Chétopodes est une larve nageuse, de forme à peu près ovoïde et divisée en deux parties par une couronne équatoriale ciliée, appelée architroque. La partie supérieure ou prostomium présente, vers son sommet, un épaississement (Scheutelplatte des zoologistes allemands); c'est le rudiment du système nerveux central, souvent surmonté d'un bouquet de cellules sensorielles ciliées. La bouche est située immédiatement au-dessous de l'architroque, dans le haut de la région du tronc (metastomium). Sa position détermine la face ventrale; le tube digestif est court et se termine par l'anus, situé vers l'extrémité inférieure du tronc, du côté de la face dorsale. L'architroque se dédouble généralement en deux couronnes cellulaires ciliées; l'une préorale, c'est l'architroque proprement dite ou organe locomoteur : l'autre, postorale, sert surtout à porter les aliments vers la bouche. Le tube digestif est formé de trois parties : un stomadoedum, un intestin moyen (archenteron) et un intestin postérieur ou prostodaeum; l'intestin moyen est endodermique, le stomadaeum et le proctodaeum sont exodermiques. La région prestomiale tout entière et la partie de la région métastomiale, comprise entre la paroi du corps et le tube digestif, forment une vaste cavité (cavité de segmentation ou cavité du corps primitive). Deux faisceaux musculaires longitudinaux, l'un dorsal, l'autre ventral, traversent le prostomium. Au-dessous de l'archétroque, de chaque côté de la bouche, on découvre des organes très intéressants : les protonèphres ou organes excréteurs primordiaux, appelés d'une façon très impropre reins céphaliques. 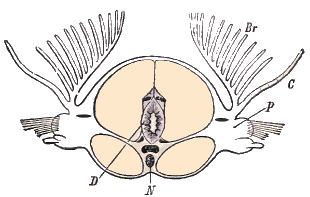
Coupe transversale d'un segment d'Eunice. D. Tube digestif et mésentères médians; N. Chaîne nerveuse et, au-dessus d'elle, vaisseau ventral de chaque côté chambres néphridiennes; P. Parapode; C, Cirre; Br. Branchies. Ces organes se trouvent dans la région antérieure du corps et non dans la tête; ils débouchent dans la cavité du corps par des entonnoirs munis de cellules flagellées (flammes vibratiles), et s'ouvrent à l'extérieur par des pores ventraux; les protonèphres de la trochosphère sont homologues des canaux excréteurs des Vers plats, des Rotifères et des Dinophitus. Dans la région du tronc se trouvent les deux bandelettes mésodermiques (Mesodermstreifen) provenant des deux cellules mésodermiques primitives qui, chez toutes les trochosphères des Chétopodes, remplacent par coenogénie les entérocoeles mésodermiques des Chaetognathes, des Brachiopodes, etc. Cette forme larvaire trochosphaere se modifie naturellement dans les divers types de Chétopodes. La larve que nous venons de décrire
est dite manotroque ou céphalotroque. On appelle telotroques
celles qui présentent, outre I'architroque, une couronne ciliaire
périanale; polytroques, celles qui présentent plusieurs
couronnes ciliaires complètes; nototroques, celles qui n'ont
que des demi-couronnes dorsales; gasterotroques, celles qui ont,
au contraire, que des demi-couronnes ventrales; amphitroques, celles
qui possèdent des demi-couronnes dorsales, alternant avec des demi-couronnes
ventrales; mésotroques, celles qui n'ont que les couronnes
intermédiaires et ont perdu l'architroque et la couronne périanale
(larves des Chaetaptères); enfin, un certain nombre de larves sont
atroques.
Chez les embryons d'Oligochètes, dont
le développement est fortement condensé, les couronnes ciliaires
ont généralement disparu et la forme trochosphère
est difficile à reconnaître.
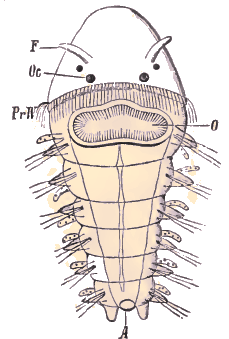
Embryon céphalotroque de Nereis. F, antennes; Oc, yeux; O, bouche; A, anus. (D'après Busch).. Régions du corps et appendicesLa valeur des parties extérieures du corps des Chétopodes a donné lieu à bien des discussions. Lorsqu'on lit les descriptions des auteurs, il est souvent difficile de savoir en particulier ce qu'ils entendent par les mots tête ou région céphalique d'une Annélide. Pour nous, le nom de tête doit être réservé à la partie qui dérive de la région prostomiale de la trochosphère. Le segment buccal, qui est le premier segment de la région métastomiale et se trouve immédiatement au-dessous de l'architroque, ne doit donc pas être compris dans la tête.- 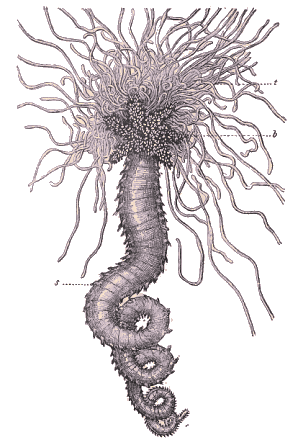
Amphitrite edwardsii. - t. Filaments tentaculaires; b. Branchies; s. Pharètres ou parapodes dorsaux. (D'après Quatrefages). Hatschek, reprenant une opinion auparavant défendue par Rathke et par Quatrefages, considérait le lobe céphalique (prostomium) et le segment buccal comme formant ensemble la tête du Polygordius. En se basant uniquement sur l'anatomie, Grube et Claparède avaient déjà remarqué l'étroite ressemblance de l'anneau buccal avec les anneaux suivants. L'embryogénie confirme absolument les données anatomiques, Dans certains cas, le prostomium avorte presque complètement chez l'adulte (nombreuses Oligochètes, etc.); les ganglions sous oesophagiens paraissent alors faire partie de l'anneau buccal. D'autres fois, au contraire, le prostomium prend un développement considérable; c'est ainsi qu'il forme les nombreux tentacules préhensiles des Térébelles et les organes appelés branchies céphaliques chez les Serpules, les Sabelles, etc. Ces organes dérivent de lobes prostomiaux homologues des vela des larves de Mollusques. Chez la plupart des Chétopodes errantes, ces lobes prostomiaux sont réduits à de courts appendices auxquels il convient de donner le nom de palpes. Pruvot a montré que les divers organes susnommés sont innervés par les nerfs stromato-gastriques. L'embryogénie ne laisse d'ailleurs aucun doute sur leur homologie. Cela a conduit à abandonner les noms d'antennes latérales, donnés aux palpes des Lycoridiens par Audouin et Milne-Edwards; les lobes frontaux des Syllidiens ne sont aussi que des palpes. On doit, au contraire, garder le nom d'antennes pour les appendices qui naissent du côté dorsal du prostomium et sont innervés par les ganglions cérébroïdes proprement dits. Ces appendices sont généralement au nombre de deux ou de trois, parfois même de cinq (certains Enniciens). Les appendices, situés de chaque côté de la tête et innervés par la chaîne nerveuse ventrale, sont des cirres plus ou moins modifiés (cirres tentaculaires) et appartiennent à l'anneau buccal et à la région métastomiale. Si l'on observe le développement
d'une trochosphère de Chétopode, on voit qu'une des premières
modifications de l'organisme larvaire consiste dans l'apparition simultanée
de trois métamères. Ce n'est qu'après un temps de
repos souvent assez long que de nouveaux segments se développent
à la suite des trois premiers avec lesquels ils présentent
parfois de nombreuses dissemblances. La portion du metastomium, qui comprend
ces trois métamères, est ce que qu'on a appelé l'archipodium;
les modifications de l'archipodium sont très intéressantes
à suivre dans les divers groupes de Chétopodes et fournissent
des indications de haute valeur pour la morphologie et la taxonomie de
ces animaux. Chez un grand nombre d'Annélides, l'archipodium demeure
formé des trois segments primaires, et tous les anneaux qui suivent
constituent la région abdominale dont les nouveaux segments se forment
d'arrière en avant, entre le dernier métamère
où pygidium et l'avant-dernier. Mais chez plusieurs Chétopodes
(Salmacina, etc.), l'archipodium continue à former de nouveaux segments,
tantôt semblables à ceux de l'archipodium primaire (Salmacina),
tantôt dissemblables (Telepsavus, etc.), mais différents également
des métamères abdominaux. L'ensemble des anneaux de l'archipodium
primaire et de la région ou des régions qui en dérivent,
est ce que les auteurs ont appelé thorax ou partie thoracique du
corps des Annélides.
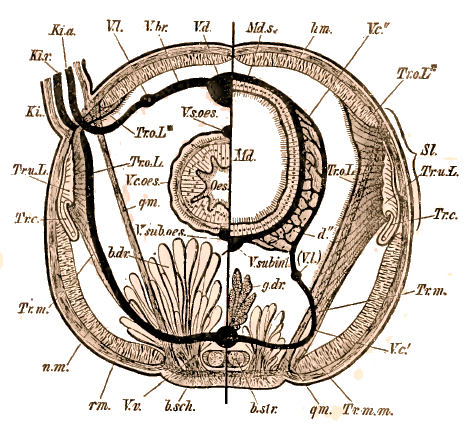
Un fait très important à signaler est que, dans toute cette partie du corps, les organes segmentaires, lorsqu'ils existent, n'ont plus en général qu'une fonction purement excrétrice. Les organes génitaux n'apparaissent que dans les segments abdominaux. Chez les Oligochètes, l'archipodium correspond le plus souvent à la région anteclitellienne. Les segments de l'archipodium primaire entrent souvent en coalescence ou sont fortement réduits et représentés seulement par les cirres tentaculaires ou des bouquets de soies rudimentaires. L'étude des Hésioniens comparés aux Neréiliens, aux Phyllodociens et aux Polynoïdiciens, permet de se rendre compte assez facilement de ces transformations. Les segments des Chétopodes sont, en général, munis de pieds (parapodes) divisés en deux rames (rame dorsale ou notopode, rame ventrale ou neuropode), et armés de soies qui servent à la locomotion et qui concourent aussi, par leurs appendices variés (branchies, cirres, etc.), à la fonction respiratoire. Les soies placées sur les pieds présentent des formes excessivement variées et fournissent de bons caractères pour la systématique. Elles sont filiformes, recourbées, aplaties, en forme de faux, de flèches, d'aiguilles, de couteaux, etc. Les pieds peuvent manquer et les soies sont alors implantées dans des invaginations de la peau (Oligochètes, Opheliadées, etc.), les soies même peuvent faire complètement défaut (Anachaeta chez les Oligochètes, Linotrypane chez les Polychètes). Dans ce cas, on trouve encore des follicules cutanés à la place où devraient se trouver les soies. D'autres fois, au contraire, les soies se multiplient au point de former sur la peau un feutrage de poils à aspect métallique et brillant sur la surface dorsale (Aphrodite). Les appendices des pieds présentent aussi une variété de formes étonnante et different parfois dans les différentes régions du corps d'un même animal, comme nous l'avons vu ci-dessus. Ce sont d'abord des filaments tentaculiformes, simples ou articulés, les cirres que l'on distingue en cirres dorsaux ou cirres ventraux. Dans d'autres cas, les cirres sont remplacés par des appendices aplatis (élytres) qui recouvrent le dos en s'imbriquant les uns sur les autres. A côté des cirres, on trouve souvent des branchies filiformes, ramifiées en touffe, en forme de peigne, etc., tantôt limitées à la région médiane du corps (Arénicole), tantôt existant sur toute la face dorsale ou seulement sur les anneaux antérieurs (Térébelles). Téguments et appareil musculaireLes téguments sont composés de deux couches, l'une externe non cellulaire, le cuticule, l'autre interne et cellulaire (corium Rthk, derme de Qtrfg., hypoderme d'un grand nombre d'auteurs). La cuticule est tantôt mince, tantôt épaisse. Partout où elle atteint une certaine épaisseur elle se montre ornée de deux systèmes de stries à angle droit ou plus souvent à 70° environ. Ces stries produisent les irisations si brillantes chez certaines espèces. Les pores tubulaires (Porencanaele des Allemands) sont distribués en lignes le long de ces stries, Claparède en a distingué deux catégories :1° ceux qui servent au déversement de certaines sécrétions; ils paraissent exister chez toutes les Annélides;L'hypoderme est en réalité le résultat de la transformation de l'exoderme embryonnaire. Il secrète la cuticule d'où le nom de couche matrice qu'on lui donne aussi quelquefois. C'est un véritable épithélium, mais il arrive souvent que chez l'animal adulte on ne distingue plus les contours des cellules. Les noyaux paraissent alors semés avec une certaine régularité dans une couche granuleuse continue; c'est ce qu'on a parfois appelé un syncytum. Parmi les glandes tégumentaires débouchant dans les pores cutanés, les unes secrètent un liquide épais, d'autres produisent des faisceaux, des bâtonnets qui ont été souvent comparés aux bâtonnets des Turbellariés, aux organes urticants des Nudibranches et des Coelentérés. Certaines familles ont leurs téguments littéralement bourrés de follicules bacillipares même dans les cirres et les antennes. C'est le cas surtout pour les Spioïdiens, les Ariciens, les Chétopteriens et un grand nombre d'Hésioniens et de Phyllodociens. La couleur des téguments est due à des amas de granulations pigmentaires dans la partie inférieure de la membrane chitinisée et aussi à des cellules de pigment situées au-dessous. Les muscles cutanés se composent d'une couche de fibres annulaires presque continue et d'une couche interne de fibres longitudiniales souvent groupés en quatre bandes; deux dorsales et deux ventrales. La structure des muscles, étudiée par Jordan et par Haswell, est souvent très élevée et rappelle celle des muscles striés des Vertébrés à leur période embryonnaire. Tube digestifLe canal digestif s'étend le plus souvent en droite ligne depuis la bouche jusqu'à l'anus situé à l'extrémité du corps du côté dorsal. Il se divise en oesophage et en intestin. A la bouche ventrale (plus rarement terminale) fait suite un pharynx souvent plus oumoins exsertile dont l'épithélium est parfois cilié mais le plus souvent cuticularisé et garni de papilles ou de denticules. L'intestin est divisé par des étranglements réguliers et une série de chambres qui correspondent aux segments extérieurs. A la partie antérieure il existe souvent une paire de culs-de-sac d'aspect glandulaire dans lesquels on trouve de l'air emmagasiné (poumons rudimentaires?). La surface extérieure de l'intestin est tapissée par des cellules mésodermiques remplies d'une substance verdâtre (cellules chloragogènes), qui paraissent jouer le rôle d'éléments de réserve pour le bourgeonnement des nouveaux individus ou des nouveaux segments. Chez quelques Annélides vivant dans le sable (Capitellides), il existe un tube intestinal accessoire analogue à celui des Spatangues. - 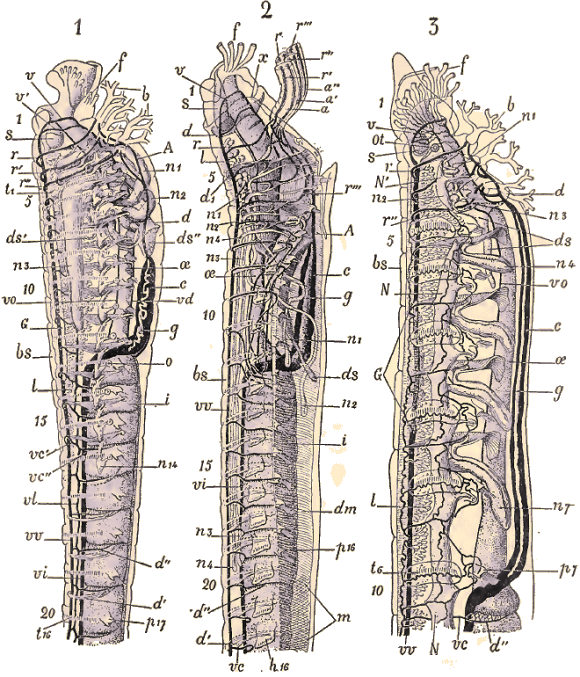
Système circulatoire et respiratoireLe système circulatoire présente un développement très variable même à l'intérieur d'une famille et parfois d'un genre déterminé. Il paraît être entièrement clos de telle sorte que le liquide nourricier qui se trouve dans la cavité générale et qui contient comme le sang des animaux supérieurs des éléments figurés, ne communique pas avec le liquide renfermé dans les vaisseaux et généralement coloré en rouge ou en vert par l'hémoglobine. La partie essentielle de l'appareil circulatoire est le vaisseau dorsal qui repose sur l'intestin dans toute son étendue et qui communique par des rameaux latéraux avec un vaisseau ventral. La circulation du sang est produite par les constrictions de certaines portions des vaisseaux et particulièrement du vaisseau dorsal contractile dans toute sa longueur ou seulement dans une portion limitée (coeur). Certaines branches latérales et plus rarement le vaisseau ventral peuvent aussi présenter des pulsations.Le sang se meut dans le vaisseau dorsal d'arrière en avant et se répand dans les anses latérales puis par des réseaux périphériques plus ou moins compliqués dans la peau et les appendices branchiaux. De ces organes le sang revient par de nouvelles anses latérales au vaisseau ventral qui se déverse à l'extrémité postérieure dans le vaisseau dorsal. Les branchies, quelles que soient leur forme et leur position, sont parcourues par un vaisseau efférent et un vaisseau afférent; les vaisseaux afférents viennent du tronc dorsal, les vaisseaux efférents se rendent au tronc ventral. Les branchies archipodiales des Térébelles, appelées parfois improprement branchies céphaliques, déterminent une modification spéciale, le vaisseau dorsal s'élargit au-dessus de l'oesophage et forme une sorte de coeur branchial qui envoie dans les branchies des rameaux pairs pendant que deux anastomoses transverses fonctionnent en même temps comme des coeurs. Les troncs longitudinaux peuvent aussi éprouver des modifications par le grand développement des ramifications et se résoudre en plexus vasculaires. Ainsi, chez Palyophthalmus le vaisseau dorsal n'existe point sur la portion moyenne du tube digestif de même que chez les Hermelles. Dans ce même point le vaisseau ventral est représenté par deux troncs. Un fait très curieux à signaler est le développement inégal du système circulatoire dans des familles voisines et parfois chez les divers représentants d'une même famille; les Aphroditiens et les Terebelliens fournissent des exemples très nets à cet égard. A côté d'espèces presque anangiées on en trouve qui sont pourvus de vaisseaux nombreux et compliqués. Il n'existe pas d'organes de respiration spéciaux chez les Oligochètes; la respiration s'accomplit par toute la surface, du tégument ou se localise dans certaines régions (Lumbriculus). Chez les Polychètes il existe des branchies dont la valeur morphologique varie avec les différents types. Tantôt ce sont des cirres dépendant des parapodes qui peuvent être très allongés (Cirratulus) ou disposés en arborescences (Amphinome) ou encore en forme de peignes (Eunice). Parfois ces cirres branchiaux peuvent se séparer des pieds et naître directement. sur la surface dorsale, soit sur la région moyenne du corps (Arenicola), soit sur tous les anneaux, mais dans ce dernier cas ils se simplifient à mesure qu'ils approchent de l'extrémité postérieure (Eunice, Amphinome). Chez les Tubicoles il existe généralement des branchies archipodiales soit sur les deux premiers téguments (Pectinaria, Sabellides), soit sur les trois premiers segments (Terebella), mais il y a de plus chez ces animaux un appareil respiratoire prostomial plus ou moins compliqué et servant en outre à d'autres fonctions (filaments préhensiles des Pectinaires et des Térébelles, bras operculaires des Spirorbes, etc.). Ces derniers ne renferment parfois que le liquide de la cavité générale (Pectinaria, Terebella), mais dans d'autres cas ils contiennent des vaisseaux sanguins (Chlorhaemiens). Chez les Sabellides ils sont soutenus par un squelette cartilagineux et munis de branches secondaires pennées. Tantôt ces filaments forment un cercle autour de la bouche, rappelant la lophophore des Bryozoaires et de Phoronis; tantôt ils forment de chaque côté un éventail (Serpulides) dont la base est souvent contournée en spirale (Spirographis). Organes d'excrétion et de sécrétionLes organes excréteurs protonéphriques que nous avons signalés chez la trochosphère des Chétopodes n'ont, lorsqu'ils existent, qu'une durée très courte. Ils sont rapidement remplacés par des deutonéphres ou organes segmentaires (Nephridies des naturalistes anglais). Signalés d'abord par Williams, ces organes ont été étudiés ensuite avec le plus grand soin par E. Meyer. Ils sont situés par paires dans tous les segments, plus rarement dans certains segments seulement (Oligochètes, Térébelliens). Leur orifice interne est infundibuliforme, cilié et libre dans la cavité coelomatique, leurs parois sont glandulaires au moins dans une portion de leur étendue. Ils sont plusieurs fois repliés sur eux-mêmes et débouchent de chaque côté du corps par un pore latéral situé dans l'anneau qui suit immédiatement celui qui contient l'infundibulum ou pavillon vibratile. Une partie de ces organes, ceux qui sont contenus dans l'archipodium primaire et secondaire, fonctionnent uniquement comme organes d'excrétion, les autres servent aussi, comme l'ont montré d'abord Ehlers et Claparède, à l'expulsion des produits génitaux. Ce sont des oviductes et des canaux déférents.Chez la plupart des Chétopodes, il existe des glandes cutanées sécrétant une substance gélatineuse destinée à englober la ponte. Chez les Oligochètes ces glandes sont localisées dans une région spéciale (clitellum), leur produit peut se coaguler dans l'eau et former une substance résistante (cocon). Beaucoup de Serpulides possèdent sur la face dorsale de la partie antérieure du corps deux grosses glandes destinées à secréter le tube qui abrite ces animaux. Chez le Siphonostonum on trouve de chaque côté de la bouche les ouvertures de deux grosses glandes renfermant des concrétions blanches. Des formations analogues existent, d'après Claparède, chez les Ammochares. Système nerveuxLe système nerveux des Chétopodes a fait l'objet d'études soignées de la part de Quatrefages, de Mac Intosh, et plus tard de Pruvot. Les ganglions cérébraux sont le plus souvent divisés en lobes très rapprochés les uns des autres et parfois entièrement confondus (Enchytraeus). Les cordons longitudinaux de la chaîne ventrale sont parfois si rapprochés l'un de l'autre qu'ils semblent ne former qu'un seul cordon (Oligochètes, Néréides, etc.). Chez les Tubicoles, au contraire, ils s'éloignent notablement l'un de l'autre de telle sorte qu'ils sont unis par des commissures souvent assez larges (Serpulides) rattachant les ganglions de chaque côté du corps. Le système stomato-gastrique se compose de ganglions pairs et impairs innervant la bouche et la trompe exsertile.Les organes des sens les plus répandus
sont les yeux dont le développement obtient
une perfection extraordinaire chez les Alciope. Généralement
situés sur le prostomium, ces organes peuvent se rencontrer parfois
à l'extrémité postérieure du corps (Fabricia),
ou affecter une disposition métamérique (Polyopthalmus).
Les filaments branchiaux des Sabelles présentent aussi des taches
pigmentaires munies de corps réfringents. Des otocystes existent
dans le voisinage du collier oesophagien chez les Arenicola, les Fabricia,
quelques Sabellides et quelques Terebellides. Les organes du tact
sont représentés par les palpes, les antennes, les cirres
et les élytres; trois organes pourvus de très riches terminaisons
nerveuses étudiées par Jourdan de Marseille. De chaque côté
du corps on observe des organes latéraux qui ont été
considérés par certains zoologistes comme des papilles gustatives.
Reproduction gemmipareLes Chétopodes peuvent reproduire les parties du corps qu'elles ont perdues soit par accident, soit par autotomie protectrice. Cette faculté de régénération s'étend même à la partie céphalique et il n'est pas rare de rencontrer des individus présentant une portion antérieure et une portion postérieure beaucoup plus jeunes que la partie moyenne du corps. La reproduction asexuée ou autotomie génératrice s'observe chez quelques Oligochètes (Aeolosama, Chaetogastridae, Naidamorphae) et chez un petit nombre de Polychètes (certains Serpuliens et certains Syllidiens). Chez Sillis prolifera une simple scission détermine la séparation d'une série d'anneaux postérieurs remplis de produits sexuels après qu'il s'est formé aux dépens d'un anneau métastomiel une tête pourvue d'yeux et de tentacules. D'autres fois et plus fréquemment ce n'est qu'un seul anneau, le dernier qui devient le point de départ de la formation d'un nouvel individu. C'est ce qu'on observe chez Autolytus prolifer. Une série d'anneaux se produisent à l'extrémité postérieure d'un oozoïde asexué et forment des chaînes soit d'individus femelles (Sacconereis helgolandica), soit d'individus mâles (Polybostrichus Mülleri). Il en est de même chez le Myrianide où le phénomène se répétant plusieurs fois entre le dernier anneau de l'individu souche et la tête de l'individu fille, il naît de la sorte une chaîne de plusieurs blastozoïtes qui après leur séparation représentent des vers sexués.- 
Reproduction sexuéeA part les Oligochètes et un petit nombre de Polychètes (quelques Néréides et quelques Serpulides) les Chétopodes sont unisexués. Les particularités de la reproduction sont particulièrement complexes chez les Néréidiens ou les formes asexuées (atoques) et les formes sexuées (épitoques) présentent parfois des dissemblances très considérables. Nous décrirons en parlant des Oligochètes l'appareil sexuel très différencié de ces Annélides. Chez le plus grand nombre des Polychètes les oeufs et les spermatozoïdes naissent de la paroi somatopleurale des dissépiments et les produits généraux à leur maturité tombent directement dans le coelome d'où ils sont expulsés par les organes segmentaires. Un petit nombre d'espèces incubent leurs oeufs à l'intérieur de l'organisme maternel et sont vivipares (Syllis vivipara). D'autres les incubent entre les tentacules céphaliques (Salmecina) ou dans un de ces tentacules transformé en organe incubateur (Spirorbis). En général, les oeufs sont déposés dans des masses gélatineuses pédonculées. Le développement est condensé chez les Oligochètes. Chez les Polychètes il y a généralement une segmentation totale mais inégale aboutissant à la formation d'une amphigastrula. Le blastopore se ferme tantôt dans le voisinage du point où se formera l'anus (Salmacina), tantôt près de la future ouverture buccale.EthologieLe genre d'existence des Chétopodes est extrêmement varié. La plupart se trouvent dans la mer; cependant les Ogolichètes sont le plus souvent d'eau douce ou terrestres. Le plus grand nombre vivent sous les pierres, dans la vase ou rampent à la surface des fonds, mais d'autres sont nageuses, pélagiques et vivent en haute mer, d'autres enfin sont parasites soit comme les Alciopides et certains Amphinomiens dans des animaux pélagiques (Cténophores, Cirripèdes), soit comme divers Euniciens chez des Gephyriens ou des Chétopodes d'autres espèces. Les Chétopodes errants sont généralement des animaux carnivores, les Tubicoles ont au contraire un régime herbivore et se nourrissent de Diatomées de débris végétaux, etc.PhosphorescencePanceri a fait une étude particulière de la phosphorescence des Annélides. Le fait était déjà connu depuis longtemps. Will l'avait signalé dès 1844 chez le Chaetopterus variopedatus Renier. Ray-Lankester a observé également la phosphorescence chez la même Annélide. Toutes les espèces du groupe ne sont cependant pas douées de cette propriété. Panceri remarque que Phyllochaetopterus socialis, espèce abondante dans le golfe de Naples, n'est jamais phosphorescent. Grube et plus tard Eiders ont signalé la phosphorescence chez Polycirrus aurantiacus et P. medusa Grube. Les espèces du genre Odontosyllis sont particulièrement remarquables, l'Odontosyllis fulgurans, peut émettre une lueur assez vive pour être perçue en plein jour, même à travers le tube qu'elle habite.Panceri a décrit aussi des Polynoïdes phosphorescentes, Polynae turcica et Pholae brevis. Une belle Polynae commensale de Synaptes, sur les côtes de Bretagne, émet également une superbe lueur d'un bleu verdâtre opalescent. La phosphorescence de Lombrics a été signalée depuis bien des années. Chez la plupart des espèces elle paraît accidentelle et temporaire. Elle est au contraire constante et très intense chez le Photodritus phosphoreus Duges. Valeur économique; usagesSur toutes les côtes de France, de Belgique, de la Grande-Bretagne, etc, l'Arenicola marina, vulgairement le ver, est employé comme amorce pour les merlans et presque toutes les sortes de poissons. On les récolte à marée basse avec une bêche à fer court et étroit. On en fait même sécher après avoir enlevé en partie le sable humide qu'elles renferment. Sur certaines plages on utilise Nephthys caeca, gravette blanche, et Nereilepas fucata. Les Néréides fournissent aussi d'excellents appâts, Nereis cultrifera et N. diversicolor. Cette dernière (carplue ou crapelure des pêcheurs du Boulonnais) se trouve particulièrement dans la vase des ports et des estuaires où l'eau douce se mêle à l'eau de mer; c'est une excellente amorce pour le bar (Labrax lupus), le mulet, etc., pour pêche à la ligne volante. On les pêche avec un fer en forme de lance et on les met dans des réceptacles contenant du sable et des herbes marines. La grande Marphyse (Marphysa sanguinea) est très estimée en Bretagne pour la pêche du congre. Si l'Annélide se brise on ne garde que la partie antérieure, le reste étant sujet à pourrir, ce qui amènerait la mort de toute la récolte. Les habitants des îles Fidji font leur régal d'une espèce voisine de Lysidice ninetta dont ils prévoient l'apparition annuelle dans leurs mers en observant les phases de la lune. Cette Annélide est appelée Palolo par les habitants des îles Samoa et Tonga, et Mbalolo par les Fidjiens. Lorsqu'elle arrive en grande quantité, les chefs les plus fortunés en expédient en présents à ceux dont les domaines ne sont pas visités par cette précieuse Annélide. Il est probable que cette Lysidice quitte ses retraites dans les rochers et récifs coralliens et se montre à la surface au moment de la reproduction. (A. Giard).PaléontologieLes Vers Chétopodes, et particulièrement ceux qui vivent dans des tubes (Tubicoles), ont laissé à l'état fossile des traces certaines de leur existence, consistant simplement dans ces tubes que l'animal habitait pendant sa vie. Les genres Serpula, Spirorbis, Terebella, Ditrupa, sont connus depuis l'époque silurienne; le genre Genicularia (Quenst.), du Jurassique supérieur, est éteint; les autres vivent encore. On trouve ces tubes fixés en parasites sur les coquilles des Brachiopodes et des Pélécypodes, les Belemnites, les Eponges, les Coraux, etc. Les Serpules abondent surtout dans le Crétacé inférieur et constituent même, dans certaines parties de l'Allemagne, de véritables bancs (Serpuliten-Kalk) formés presque exclusivement de ces tubes (Serpula coacervata, S. gordialis).La distinction des genres vivants reposant essentiellement sur l'organisation de l'animal et la forme de l'opercule, qui ne se conservent pas à l'état fossile, on conçoit que la classification des formes éteintes doive rester forcément dans le vague; pour cette même raison, la création des genres fossiles, proposés par différents auteurs, n'est pas suffisamment justifiée. Certaines formes peuvent être confondues avec les coquilles tubuleuses du genre Vermetus, mais s'en distinguent par l'absence de cloisons internes; d'autres (Ditrupa) ressemblent aux coquilles de Dentale, mais se reconnaissent toujours à ce que l'une des extrémités du tube est toujours fermée chez les Serpules. (E. Trt). |
| . |
|
|
|||||||||||||||||||||||||||||||
|