 |
La cellule
est la plus petite unité d'un organisme vivant; elle en est l'élément
constitutif de base. Les organismes les plus simples (Bactéries, Protistes,
etc.) sont composés d'une seule cellule. Dans les organismes pluricellulaires
(Animaux, Plantes, etc.), les cellules coopÚrent pour assurer les différentes
fonctions vitales de l'organisme auquel elles appartiennent, et selon la
fonction qui leur revient, elles revĂȘtent des caractĂ©ristiques
spéciales. Chez les Animaux, par exemple, les cellules osseuses aident
à soutenir et à protéger le corps. Les cellules du systÚme immunitaire
combattent les Bactéries envahissantes. De plus, les globules rouges transportent
l'oxygĂšne dans tout l'organisme. Chacun de ces types de cellules joue
un rÎle vital pendant la croissance, le développement et l'entretien
quotidien de l'organisme. Les cellules d'un mĂȘme type s'interconnectent
les unes aux autres et remplissent des fonctions communes pour former des
tissus (par exemple, le tissu musculaire, le tissu conjonctif et le tissu
nerveux), plusieurs tissus se combinent pour former un organe (par exemple,
l'estomac, le coeur ou le cerveau), et plusieurs organes constituent un
systĂšme organique (comme le systĂšme digestif, le systĂšme circulatoire
ou le systĂšme nerveux). Au final, ce sont donc tous ces systĂšmes, qui
fonctionnant ensemble, définissent l'organisme (comme un éléphant, par
exemple). Malgré leur énorme variété, toutes les cellules partagent
cependant certaines caractéristiques fondamentales.
-
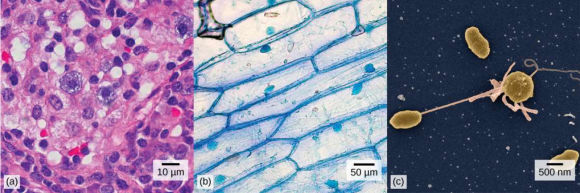
| (a)
Les
cellules des sinus nasaux (vues avec un microscope optique), (b) les
cellules d'oignons (vues avec un microscope optique), et (c) les
cellules bactériennes Vibrio tasmaniensis (vues avec un microscope
électronique à balayage) proviennent d'organismes trÚs différents,
mais tous partagent certaines caractéristiques de la structure cellulaire
de base. (crédits : a) d'Ed Uthman, MD; b)
Umberto Salvagnin; c) Anthony D'Onofrio; données d'échelle de Matt Russell). |
Cellules procaryotes
et eucaryotes
On distingue deux grandes catégories de cellules
: les cellules procaryotes et les cellules eucaryotes.
Les cellules procaryotes (pro-
= avant; -karyon- = noyau) n'ont pas de noyau; leur matériel génétique
se concentre dans une région de la cellule appelée nucléoïde. Les cellules
eucaryotes (eu- = vrai) possÚdent au contraire un noyau bien délimité,
entouré par une membrane, qui renferme leur matériel génétique.
 On
défini les deux plus grandes divisions du vivant en substantivant les
adjectifs eucaryote et procaryote. Les Procaryotes sont les organismes
formés de cellules procaryotes : il s'agit des organismes à prédominance
unicellulaire qui forment les domaines des
Bactéries
et des Archées. Les
Eucaryotes sont les organismes formés de cellules
eucaryotes : il s'agit des Animaux, des Plantes,
des Champignons, des Algues
et des Protistes. On
défini les deux plus grandes divisions du vivant en substantivant les
adjectifs eucaryote et procaryote. Les Procaryotes sont les organismes
formés de cellules procaryotes : il s'agit des organismes à prédominance
unicellulaire qui forment les domaines des
Bactéries
et des Archées. Les
Eucaryotes sont les organismes formés de cellules
eucaryotes : il s'agit des Animaux, des Plantes,
des Champignons, des Algues
et des Protistes.
Quelle que soit leur catégorie, toutes les
cellules partagent quatre composants communs :
1) une membrane plasmique (qu'on
appelait autrefois la membrane cellulaire), qui est un revĂȘtement extĂ©rieur
séparant l'intérieur de la cellule de son environnement;
2) un cytoplasme, une substance gélatineuse
à l'intérieur de la cellule dans laquelle se trouvent d'autres composants
cellulaires;
3) de l'ADN
(acide désoxyribonucléique), qui est le matériel génétique de la cellule;
4) des ribosomes, qui sont les particules
qui synthétisent les protéines.
Procaryotes ou eucaryotes,
toutes les cellules présentent une grande diversité de formes et de tailles,
qui résultent de leur adaptation évolutive à différents environnements
ou à différentes fonctions spécialisées au sein des organismes pluricellulaires.
-
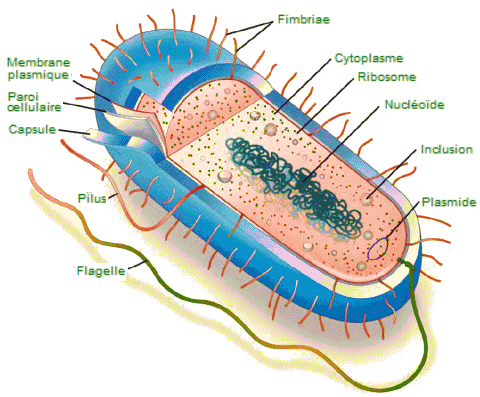
| Structure
générale d'une cellule procaryote. - Une cellule procaryote typique
contient une membrane cellulaire, de l'ADN chromosomique concentré dans
un nucléoïde, des ribosomes et une paroi cellulaire. Certaines cellules
procaryotes peuvent également posséder des flagelles, des pili (poils),
des fimbriae (franges) et des capsules. |
Taille des cellules.
Avec de dimensions de l'ordre de 0,1 Ă
5,0 ”m, les cellules procaryotes sont significativement plus petites que
les cellules eucaryotes, dont les diamÚtres se situent généralement
entre 10 Ă 100 ”m (certaines pouvant mĂȘme ĂȘtre beaucoup plus grandes,
comme les oeufs des Oiseaux).
La petite taille des cellules procaryotes
permet aux ions et aux molécules organiques qui y pénÚtrent de se propager
rapidement Ă d'autres parties de la cellule. De mĂȘme, tous les dĂ©chets
produits dans une cellule procaryote peuvent rapidement sortir. Ce n'est
pas le cas des cellules eucaryotes dont l'Ă©volution s'est traduite par
différentes adaptations structurelles pour améliorer le transport cellulaire.
La grande taille de ces cellules n'aurait pas été possible sans ces adaptations.
En général, la taille des cellules est
limitée, car le volume (proportionnel au cube du diamÚtre) augmente beaucoup
plus rapidement que la surface cellulaire (proportionnelle au carré du
diamĂštre). Lorsqu'une cellule devient plus grande, il devient de plus
en plus difficile pour la cellule d'acquérir suffisamment de matériaux
pour soutenir les processus à l'intérieur de la cellule, car la taille
relative de la surface, autrement dit les voies de passage, Ă travers
laquelle les matĂ©riaux doivent ĂȘtre transportĂ©s diminue.
Les cellules eucaryotes
Une cellule eucaryote est une cellule qui
a son noyau lié à la membrane et d'autres compartiments ou sacs liés
à la membrane, appelés organites (= petits organes). Ceux-ci ont des
fonctions spécialisées, tout comme les organes du corps ont des fonctions
spĂ©cialisĂ©es. De mĂȘme, ces organites permettent aux diverses fonctions
qu'ils assurent de se produire simultanément dans la cellule. Les cellules
eucaryotes ont ainsi une structure plus complexe que les cellules procaryotes.
-
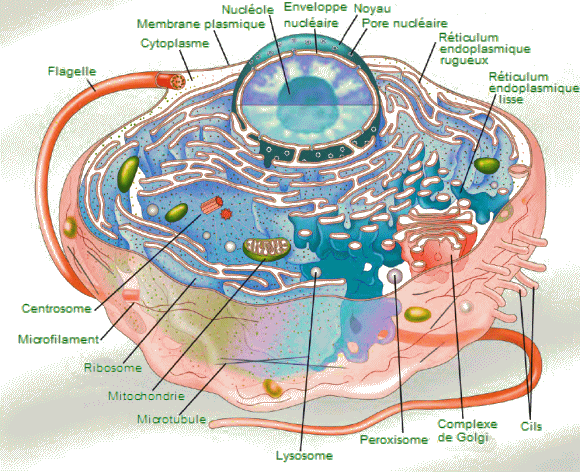
| Cellule
eucaryote d'un organisme unicellulaire. - Notez que les cellules des
Eucaryotes ont des formes et des des fonctions trĂšs variables, aussi n'y
trouve-t-on pas nécessairement toutes structure représentées ici. |
Avant de discuter plus en détail des fonctions
des organites au sein d'une cellule eucaryote, examinons d'abord deux composants
importants de la cellule : la membrane plasmique et le cytoplasme.
La membrane plasmique.
Comme les cellules procaryotes, les cellules
eucaryotes ont une membrane plasmique composée d'une bicouche phospholipidique
avec
des protéines incorporées, qui sépare le contenu de la cellule de son
environnement. Un phospholipide est une molécule
lipidique composée de deux chaßnes d'acides gras, d'un squelette
glycérol et d'un groupe phosphate. La membrane plasmique régule le passage
de certaines substances, telles que les molécules organiques, les ions
et l'eau, empĂȘchant le passage de certaines pour maintenir les conditions
internes, tout en introduisant ou en retirant activement d'autres. Certains
composés peuvent aussi se déplacer passivement à travers la membrane.
Les membranes plasmiques des cellules qui
se spécialisent dans l'absorption sont repliées pour prendre des formes
semblables à des doigts appelées microvillosités. Ce pliage augmente
la surface de la membrane plasmique. Ces cellules se trouvent généralement
le long de l'intestin grĂȘle, l'organe
qui absorbe les nutriments des aliments digérés.
Les
personnes atteintes d'une maladie coeliaque développent une réponse immunitaire
au gluten, qui est une protéine présente dans le blé,
l'orge et le
seigle. La
réponse immunitaire endommage les microvillosités et, par conséquent,
les personnes atteintes ne peuvent pas absorber les nutriments. Cela conduit
à de la malnutrition, à des crampes et à de la diarrhée. Les patients
souffrant d'une telle maladie doivent suivre un régime sans gluten.
Le cytoplasme.
Le cytoplasme correspond au contenu
d'une cellule entre la membrane plasmique et l'enveloppe nucléaire. Il
est composé de cytosol (un fluide de type gel), d'organites en suspension
dans celui-ci, du cytosquelette qui procure une armature Ă la cellule,
et de diverses substances chimiques. MĂȘme si le cytoplasme est constituĂ©
de 70% Ă 80% d'eau, il a une consistance semi-solide, qui provient des
protéines qu'il contient. Cependant, les protéines ne sont pas les seules
molécules organiques présentes dans le cytoplasme. On y trouve également
du glucose et d'autres sucres simples, des polysaccharides, des acides
aminés, des acides nucléiques, des acides gras et des dérivés du glycérol.
Des ions de sodium, de potassium, de calcium et de nombreux autres éléments
sont également dissous dans le cytoplasme. De nombreuses réactions métaboliques,
à commencer par la synthÚse des protéines, ont lieu dans le cytoplasme.
Les
organites.
Les organites
sont les structures subcellulaires que l'on rencontre dans le cytoplasme
des cellules et qui ont une forme et une fonction qui leur est propre.
Certains organites sont communs Ă toutes les cellules eucaryotes (mitochondries,
ribosomes, peroxysome, réticulum endoplasmique, vacuoles et vésicules,
appareil de Golgi), d'autres sont propres aux cellules animales (centrosome,
lysosome), et d'autres encore ne se rencontrent que dans les cellules végétales
(paroi cellulaire, chloroplastes, grande
vacuole). Les ribosomes, par exemple, sont les centres de production des
protéines, les lysosomes décomposent les
protéines,
les polysaccharides, les lipides,
les acides nuclĂ©iques et mĂȘme les organites
usés, les chloroplastes sont les organites qui assurent la photosynthÚse
par les plantes, etc.
Le
systĂšme endomembranaire.
Le systĂšme endomembranaire (endo
= intérieur) est un groupe de membranes et d'organites dans les cellules
eucaryotes qui travaillent ensemble pour modifier, conditionner et transporter
les lipides et les protéines. Il comprend l'enveloppe nucléaire, le réticulum
endoplasmique, les vésicules, l'appareil de Golgi, auxquels s'ajoutent,
dans les cellules animales, les lysosomes. Bien qu'elle ne soit pas techniquement
à l'intérieur de la cellule, la membrane plasmique est incluse dans le
systĂšme endomembranaire car elle interagit avec les autres organites de
ce systĂšme.
-
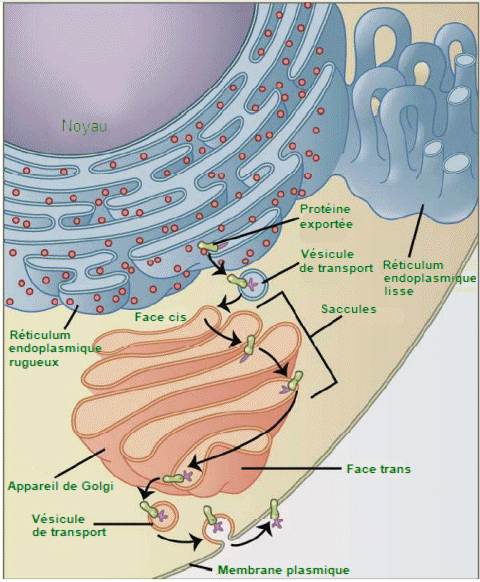
| Le
rĂŽle du systĂšme endomembranaire est de modifier, emballer et transporter
entre le réticulum endoplasmique et la membrane plasmique des lipides
et des protéines. (Crédit: Magnus Manske). |
Le
cytosquelette.
On donne le nom de cytosquelette
au rĂ©seau de fibres protĂ©iques contenu dans le cytoplasme, qui aide Ă
maintenir la forme de la cellule, sécurise certains organites dans des
positions spécifiques, permet au cytoplasme et aux vésicules de se déplacer
à l'intérieur de la cellule et permet aussi aux organismes unicellulaires
de se déplacer indépendamment.
.
Les composantes
des cellules
| Composant
cellulaire |
Fonction |
Présent
chez les Procaryotes? |
Présent
dans les cellules animales? |
Présent
dans les cellules végétales? |
|
Membrane
plasmique
|
SĂ©pare
la cellule de l'environnement extérieur; contrÎle le passage des molécules
organiques, des ions, de l'eau, de l'oxygÚne et des déchets dans et hors
de la cellule |
Oui |
Oui |
Oui |
|
Cytoplasme
|
Fournit
une structure à la cellule; site de nombreuses réactions métaboliques;
milieu dans lequel se trouvent des organites |
Oui |
Oui |
Oui |
|
Nucléoïde
|
Emplacement
de l'ADN |
Oui |
Non |
Non |
|
Noyau
|
Organite
cellulaire qui abrite l'ADN et dirige la synthĂšse des ribosomes et des
protéines |
Non |
Oui |
Oui |
|
Ribosomes
|
SynthĂšse
des protéines |
Oui |
Oui |
Oui |
|
Mitochondries
|
Production
d'ATP / respiration cellulaire |
Non |
Oui |
Oui |
|
Peroxisomes
|
Oxyde
et décompose les acides gras et les acides aminés et détoxifie les poisons |
Non |
Oui |
Oui |
VĂ©sicules
et
vacuoles
|
Stockage
et transport; fonction digestive dans les cellules végétales |
Non |
Oui |
Oui |
|
Centrosome
|
RĂŽle
non spécifié dans la division cellulaire dans les cellules animales;
centre organisateur des microtubules dans les cellules animales |
Non |
Oui |
Non |
|
Lysosomes
|
Digestion
des macromolécules; recyclage des organites usés |
Non |
Oui |
Non |
|
Paroi
cellulaire
|
Protection,
support structurel et maintien de la forme des cellules |
Oui
(principalement peptoglycane dans les bactéries, mais pas dans les Archées) |
Non |
Oui (principalement
cellulose) |
|
Chloroplastes
|
PhotosynthĂšse |
Non |
Non |
Oui |
|
RĂ©ticulum
endoplasmique
|
Modifie
les protéines et synthétise les lipides |
Non |
Oui |
Oui |
Appareil
de
Golgi
|
Modifie,
trie, étiquette, conditionne et distribue les lipides et les protéines |
Non |
Oui |
Oui |
|
Cytosquelette
|
Maintient
la forme des cellules, sécurise les organites dans des positions spécifiques,
permet au cytoplasme et aux vésicules de se déplacer à l'intérieur
de la cellule et permet aux organismes unicellulaires de se déplacer indépendamment |
Oui |
Oui |
Oui |
|
Flagelles
|
Locomotion
cellulaire |
Parfois |
Parfois |
Non,
sauf exception |
|
Cils
|
Locomotion
cellulaire, mouvement des particules le long de la surface extracellulaire
de la membrane plasmique et filtration |
Non |
Parfois |
Non |
Matrice extracellulaire
et jonctions intercellulaires.
La plupart des cellules animales libĂšrent
des matériaux dans l'espace extracellulaire. Les principaux composants
de ces matériaux sont les glycoprotéines et le collagÚne
protéique. Collectivement, ces matériaux constituent la matrice extracellulaire
ou ciment intercellulaire). Non seulement la matrice extracellulaire
maintient les cellules ensemble pour former un
tissu,
mais elle permet également aux cellules à l'intérieur du tissu de communiquer
entre elles, via des
jonctions itercellulaires
: jonctions serrées, desmosomes, jonctions lacunaires. Les cellules
végétales sont connectées et communiquent avec leurs voisines via des
plasmodesmes.
La
matrice extracellulaire.
Lorsque des récepteurs
protéiques à la surface de la membrane plasmique d'une cellule animale
se lient à une substance adaptée à la matrice extracellulaire, une chaßne
de rĂ©actions commence qui modifie les activitĂ©s qui se dĂ©roulent Ă
l'intérieur de la cellule.
La coagulation sanguine fournit un exemple
du rĂŽle que joue la matrice extracellulaire dans la communication cellulaire.
Quand les cellules tapissant un vaisseau sanguin
sont endommagées, elles présentent un récepteur protéique appelé facteur
tissulaire. Lorsque le facteur tissulaire se lie Ă un autre facteur
de la matrice extracellulaire, il fait adhérer les plaquettes à la paroi
du vaisseau sanguin endommagé, stimule les cellules musculaires lisses
adjacentes du vaisseau sanguin Ă se contracter (resserrant ainsi le vaisseau
sanguin) et initie une série d'étapes qui stimulent les plaquettes pour
produire des facteurs de coagulation.
-
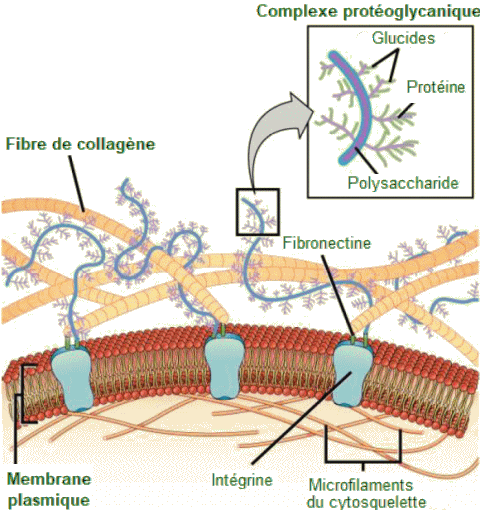
La
matrice
extracellulaire est constituée d'un réseau
de
substances sécrétées par la cellule.
Jonctions
intercellulaires.
Les jonctions intercellulaires, on l'a
dit, sont de différents types :
âą Les plasmodesmes. -
En général, de longs tronçons des membranes plasmiques des cellules
végétales voisines ne peuvent pas se toucher car ils sont séparés par
les parois cellulaires entourant chaque cellule. Les plasmodesmes sont
de nombreux canaux qui passent entre les parois cellulaires des cellules
végétales adjacentes, reliant leur cytoplasme et permettant aux molécules
de signal et aux nutriments d'ĂȘtre transportĂ©s de cellule Ă cellule.
⹠Les jonctions serrées. - On
désigne ainsi les jonctions étanches à l'eau entre deux cellules animales
adjacentes. Les protéines maintiennent étroitement les cellules les unes
contre les autres. Cette adhĂ©rence serrĂ©e empĂȘche les matĂ©riaux de
se perdre entre les cellules. Des jonctions serrées se trouvent généralement
dans le tissu épithélial qui tapisse les organes internes et les cavités,
et compose la majeure partie de la peau. Les jonctions
serrées des cellules épithéliales tapissant la vessie,
par exemple, empĂȘchent l'urine de fuir dans l'espace
extracellulaire.
âą Les desmosomes (jonctions d'ancrage).
- On trouve Ă©galement uniquement dans les cellules animales des desmosomes,
qui agissent comme des soudures par points entre les cellules épithéliales
adjacentes. Ils maintiennent les cellules ensemble sous la forme d'un feuillet
Ă©lastique dans des organes et des tissus qui s'Ă©tirent, comme la peau,
le coeur et les muscles.
âą Les jonctions lacunaires
(jonctions communicantes ou jonctions perméables). - Les jonctions
lacunaires dans les cellules animales sont
analogues aux plasmodesmes dans les cellules végétales : ce sont
des canaux entre les cellules adjacentes qui permettent le transport des
ions, des nutriments et d'autres substances qui permettent aux cellules
de communiquer. Structurellement, cependant, les jonctions lacunaires et
les plasmodesmes diffĂšrent.
Les jonctions lacunaires apparaissent lorsqu'un
ensemble de six protéines (appelées connexines) dans la membrane
plasmique s'organisent dans une configuration allongée en forme de beignet
appelée connexon. Lorsque les pores des connexons dans les cellules
animales adjacentes s'alignent, un canal entre les deux cellules se forme.
Les jonctions lacunaires sont particuliĂšrement
importantes dans le muscle cardiaque : le signal Ă©lectrique pour que le
muscle se contracte passe efficacement Ă travers les jonctions lacunaires,
permettant aux cellules du muscle cardiaque de se contracter ensemble.
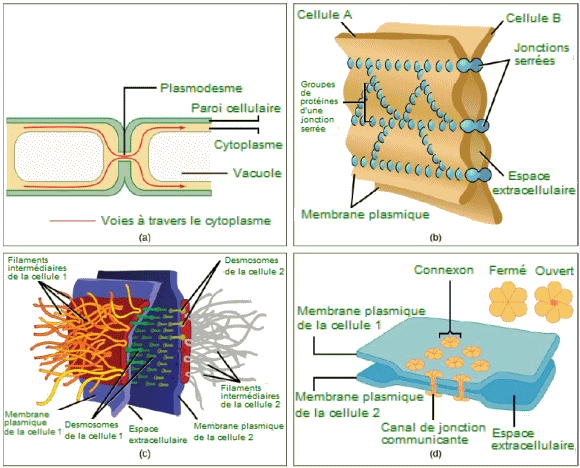
| Jonctions
cellulaires. - Il existe quatre types de connexions entre les cellules.
(a) Un plasmodesme est un canal entre les parois cellulaires de deux cellules
végétales adjacentes. (b) Des jonctions serrées joignent les cellules
animales adjacentes. (c) Les desmosomes joignent deux cellules animales
ensemble. (d) Les jonctions lacunaires agissent comme des canaux entre
les cellules animales. (crédits b, c, d :Mariana
Ruiz Villareal). |
La membrane plasmique
La membrane plasmique d'une cellule enveloppe
entiÚrement celle-ci : elle définit donc la limite de la cellule et détermine
la nature de son contact avec l'environnement. Les cellules excluent certaines
substances, en absorbent d'autres et en excrĂštent
d'autres encore, toutes en quantités contrÎlées.
La membrane plasmique
agit dÚs lors comme une barriÚre entre l'intérieur et l'extérieur de
la cellule en permettant l'entrée et la sortie de ces substances. Ce passage
est un phénomÚne appelé
perméabilité et cette perméabilité
est sélective : certaines substances doivent pouvoir entrer dans la cellule
ou en sortir, les matiĂšres nocives ne doivent pas pouvoir rentrer
et les matiĂšres essentielles de doivent pas ĂȘtre en mesure de sortir.
La membrane plasmique doit Ă©galement ĂȘtre
suffisamment flexible pour permettre Ă certaines cellules, telles que
les globules rouges et les globules blancs, de changer de forme lorsqu'elles
traversent des capillaires Ă©troits. La membrane plasmique n'est donc pas
un simple sac statique. Elle est dynamique et en constante Ă©volution.
De plus, la surface de la membrane plasmique
porte des marqueurs (molécules particuliÚres) qui permettent aux cellules
de se reconnaßtre mutuellement, ce qui est vital au cours du développement
précoce d'un organisme, à mesure que les tissus et les organes se forment.
Cette reconnaissance mutuelle joue plus tard un rĂŽle dans la distinction
entre "soi" et "non-soi", qui est le fondement de la réponse immunitaire.
La membrane plasmique porte aussi des récepteurs
qui sont des sites de fixation pour des substances spécifiques qui interagissent
avec la cellule. Chaque récepteur est structuré pour se lier à une substance
spécifique. Par exemple, les récepteurs de surface de la membrane créent
des changements à l'intérieur, comme ceux observés dans les enzymes
des voies mĂ©taboliques. Ces voies mĂ©taboliques peuvent ĂȘtre vitales
pour fournir de l'énergie à la cellule, fabriquer des substances spécifiques
pour la cellule, décomposer les déchets cellulaires ou éliminer des
toxines.
Les récepteurs situés à la surface extérieure
de la membrane plasmique interagissent avec les hormones
ou les neurotransmetteurs et permettent
Ă leurs messages d'ĂȘtre transmis Ă la cellule. Certains sites de reconnaissance
sont utilisés par les
virus comme points d'attache.
Bien qu'ils soient hautement spécifiques, des agents pathogÚnes comme
les virus peuvent évoluer pour exploiter les récepteurs pour entrer dans
une cellule en imitant la substance spécifique que le récepteur est censé
se lier. Cette spécificité permet, par exemple, d'expliquer pourquoi
le virus de l'immunodéficience humaine (VIH) ou l'un des cinq types de
virus de l'hépatite envahissent uniquement des cellules spécifiques.
Le
modĂšle de la mosaĂŻque fluide.
En 1972, S. J. Singer et Garth L. Nicolson
ont proposé un nouveau modÚle de la membrane plasmique qui, par rapport
à une compréhension antérieure, expliquait mieux les observations microscopiques
et la fonction de la membrane plasmique. Cela s'appelait le modĂšle
de la mosaïque fluide. Le modÚle a quelque peu évolué au fil du
temps, mais il rend mieux compte de la structure et des fonctions de la
membrane plasmique telles qu'on les comprend aujourd'hui.
Le modÚle de mosaïque fluide décrit
la structure de la membrane plasmique comme une mosaĂŻque de composants
- parmi lesquels des phospholipides, du cholestérol,
des protéines et des glucides - dans laquelle ces composants peuvent s'écouler
et changer de position, tout en maintenant l'intégrité de base de la
membrane. Les molécules de phospholipides et les protéines intégrées
sont capables de diffuser rapidement et latéralement dans la membrane.
-
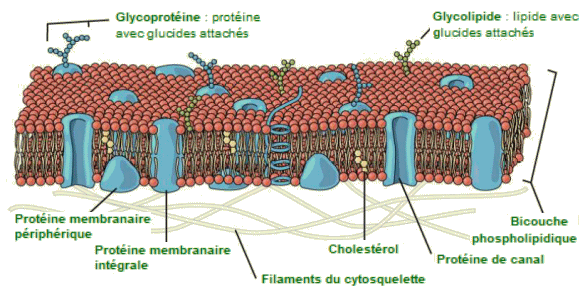
| ModĂšle
de la mosaĂŻque fluide de la membrane plasmique. - La membrane plasmique
est une bicouche phospholipidique avec des protéines incorporées. D'autres
composants, tels que le cholestĂ©rol et les glucides, peuvent ĂȘtre trouvĂ©s
dans la membrane en plus des phospholipides et des protéines. Le modÚle
de mosaïque fluide de la structure de la membrane plasmique décrit cette
membrane comme une combinaison fluide de ces diverses substances. |
La fluidité de la membrane plasmique est
nécessaire aux activités de certaines enzymes et transportent des molécules
à l'intérieur de la membrane. Les membranes plasmiques ont une épaisseur
de 5 Ă 10 nm. Ă titre de comparaison, les globules rouges humains ont
une épaisseur d'environ 8 ”m, ce qui est environ 1000 fois plus gros
que l'Ă©paisseur d'une membrane plasmique.
La membrane plasmique est constituée principalement
d'une bicouche de phospholipides dans laquelle sont incluses de nombreuses
protéines (= mosaïque protéique). Celles qui se trouvent aux surfaces
intérieure et extérieure de la bicouche sont dites protéines périphériques;
celles qui pénÚtrent à l'intérieur de la bicouche sont dites protéines
inétgrales. Toutes ces molécules (protéines et phospholipides) peuvent
ĂȘtre unies Ă des glucides pour former des glycoprotĂ©ines.et des glycolipides.
Les membranes des cellules animales peuvent
également intégrer du cholestérol.
La quantité de ce cholestérol dans les membranes plasmiques animales
régule la fluidité de la membrane et change en fonction de la température
de l'environnement cellulaire. En d'autres termes, le cholestérol agit
comme un antigel dans la membrane cellulaire et est plus abondant chez
les animaux qui vivent dans des climats froids.
âą
Phospholipides. - Le tissu principal de la membrane est composé de deux
couches de molécules de phospholipides, et les
extrémités polaires de ces molécules sont en contact avec un fluide
aqueux à l'intérieur et à l'extérieur de la cellule. Ainsi, les deux
surfaces de la membrane plasmique sont hydrophiles. En revanche, l'intérieur
de la membrane, entre ses deux surfaces, on trouve une région hydrophobe
ou non polaire Ă cause des queues d'acides gras des phospholipides. Cette
région n'a aucune attraction pour l'eau ou d'autres molécules polaires.
⹠Protéines.
- Les protéines constituent le deuxiÚme composant
chimique majeur des membranes plasmiques.
+ Les protéines
intégrales sont insérées dans la membrane plasmique et peuvent s'étendre
sur tout ou partie de la membrane. Les protéines intégrales peuvent servir
de canaux ou de pompes (canaux protéiques) pour déplacer les matériaux
dans ou hors de la cellule.
+ Les protéines
périphériques se trouvent sur les surfaces extérieures ou intérieures
des membranes, attachées soit aux protéines intégrales soit aux molécules
de phospholipides.
Les protéines intégrales
et périphériques peuvent servir d'enzymes, de points d'ancrage structuraux
pour les fibres du cytosquelette ou de sites de reconnaissance cellulaire.
âą Glucides.
- Les
glucides sont le troisiĂšme composant majeur
des membranes plasmiques. Ils se trouvent toujours à la surface extérieure
des cellules et sont liés soit aux protéines (formant des glycoprotéines)
soit aux lipides (formant des glycolipides). Ces chaĂźnes glucidiques peuvent
ĂȘtre constituĂ©es de 2 Ă 60 unitĂ©s monosaccharides et peuvent ĂȘtre
droites ou ramifiées. Avec les protéines périphériques, les glucides
forment des sites spécialisés à la surface des cellules qui permettent
aux cellules de se reconnaĂźtre.
Toutes les autres membranes
de la cellule (celles du noyau, des mitocondries, de l'appareil de Golgi,
des chloroplastes, etc.), possĂšdent une
structure similaire; seules les proportions et les types de lipides, de
protéines et de glucides pouvant différer selon l'organite considéré.
Fluidité
membranaire.
Dans le modĂšle de la mosaĂŻque fluide,
les protéines intégrales et les lipides existent dans la membrane sous
forme de molécules séparées mais reliées de maniÚre lùche. L'arrangement
de ces molécule ressemble à celui des carreaux multicolores séparés
d'une image en mosaïque, et ils flottent, se déplaçant quelque peu les
uns par rapport aux autres. La membrane n'est pas pour autant semblable
Ă un ballon qui peut se dilater et se contracter; elle est plutĂŽt assez
rigide et peut éclater si elle est pénétrée ou si la cellule absorbe
trop d'eau. En raison de sa nature mosaĂŻque, une aiguille trĂšs fine ,
cependant, peut facilement pénétrer une membrane plasmique sans la faire
Ă©clater, et la membrane se refermera automatiquement lorsque l'aiguille
sera extraite.
La structure mosaĂŻque de la membrane explique
au moins en partie sa fluidité. Un facteur qui aide à maintenir celle-ci
tient Ă la nature des phospholipides eux-mĂȘmes. Dans leur forme saturĂ©e,
les acides gras contenus dans les queues de phospholipides sont saturés
d'atomes d'hydrogĂšne
liés. Il n'y a pas de doubles liaisons entre les atomes de carbone adjacents.
Il en résulte des queues relativement droites. En revanche, les acides
gras insaturés ne contiennent pas un nombre maximal d'atomes d'hydrogÚne,
mais ils contiennent des doubles liaisons entre les atomes de carbone adjacents;
une double liaison se traduit par un coude dans la chaĂźne d'atomes de
carbone d'environ 30 degrés.
Ainsi, si les acides gras saturés, avec
leur queue droite, sont comprimés lorsque la température baisse, ils
se pressent les uns contre les autres, formant une membrane dense et assez
rigide. Si les acides gras insaturés sont comprimés, les "plis" dans
leur queue couchent les molécules de phospholipides adjacentes, en maintenant
un certain espace entre les molĂ©cules de phospholipides. Cela aide Ă
maintenir la fluidité de la membrane à des températures auxquelles les
membranes contenant des queues d'acides gras saturés dans leurs phospholipides
« gÚleraient » ou se solidifieraient.
La fluidité relative de la membrane est
particuliĂšrement importante dans un environnement froid. Un tel environnement
a tendance à comprimer les membranes composées en grande partie d'acides
gras saturés, les rendant moins fluides et plus susceptibles de se rompre.
De nombreux organismes (les Poissons en sont
un exemple) sont capables de s'adapter aux environnements froids en modifiant
la proportion d'acides gras insaturés dans leurs membranes en réponse
à la baisse de la température.
Le transport passif.
La
membrane cellulaire peut ĂȘtre permĂ©able Ă certaines substances (molĂ©cules
ou corps plus gros). Cette traversée de la membrane peut se faire principalement
selon deux modalités, le transport passif, dans lequel des molécules
(de petite taille) peuvent traverser la membrane sans apport d'Ă©nergie
par la cellule, et le transport actif (évoqué plus bas) recourt de l'énergie
produite par la cellule, cela permet le transport de petites molécules,
mais aussi de corps plus gros (phénomÚnes d'endocytose et d'exocytose).
Le transport membranaire
passif se fait généralement le long d'un gradient de concentration, c'est-à -dire
du cĂŽtĂ© oĂč une substance est plus concentrĂ©e vers le cĂŽtĂ© oĂč elle
est moins concentrée. Les molécules impliquées dans le transport membranaire
passif comprennent un large Ă©ventail de substances, notamment les ions
(comme Na+, K+,
Ca2+, Cl-), les petites molécules
non chargées (telles que l'eau, l'oxygÚne et le dioxyde de carbone),
ainsi que d'autres molĂ©cules plus grandes qui peuvent ĂȘtre spĂ©cifiquement
transportées par des transporteurs membranaires. Les principaux types
de transport membranaire passif sont :
Diffusion
simple.
Les petites molécules
non chargées, telles que l'oxygÚne (O2) et le dioxyde
de carbone (CO2), peuvent diffuser directement Ă
travers la bicouche lipidique de la membrane cellulaire en raison de leur
petite taille et de leur nature liposoluble.
Diffusion
facilitée.
La diffusion facilitée
correspond Ă la situation oĂč des molĂ©cules plus grosses ou chargĂ©es
nécessitent l'aide de protéines de transport intégrales pour traverser
la membrane. Ces protéines peuvent former des canaux (canaux
ioniques) ou des transporteurs (transporteurs
membranaires) qui facilitent le passage sélectif de certaines substances.
Par exemple, les canaux ioniques permettent le passage sélectif d'ions
tels que le sodium (Na+), le potassium
(K+), le calcium (Ca2+),
et le chlore (Cl-) Ă travers la membrane.
Osmose.
L'osmose est un
processus particulier de diffusion de l'eau à travers une membrane semi-perméable
qui se produit du cĂŽtĂ© oĂč la concentration en solutĂ©s est plus Ă©levĂ©e
vers le cĂŽtĂ© oĂč elle est plus faible. Cela Ă©quilibre la concentration
de solutés de part et d'autre de la membrane.
Le transport actif.
Le transport actif
dans les membranes cellulaires fait référence au mouvement de substances
Ă travers la membrane cellulaire contre leur gradient de concentration,
c'est-à -dire de régions de concentration plus faible vers des régions
de concentration plus élevée. Contrairement au transport passif, qui
se produit de maniÚre spontanée et ne nécessite pas d'énergie cellulaire,
le transport actif requiert de l'énergie, généralement sous forme d'ATP
(adénosine triphosphate) dans le cas des cellules vivantes. Il existe
plusieurs types de transport actif : transport par pompe ionique et, pour
les particules plus grosses, transport par endocytose et transport par
exocytose.
Transport
par pompe ionique.
Les pompes ioniques
sont des protéines membranaires spécialisées qui utilisent l'énergie
de l'ATP pour transporter des ions Ă travers les membranes cellulaires
contre leur gradient de concentration. Ces pompes permettent de maintenir
l'homéostasie ionique et de générer des potentiels électriques
Ă travers les membranes cellulaires dans de nombreux types de cellules.
Exemples :
âą La
pompe sodium-potassium (Na+/K+-ATPase) transporte trois ions sodium
(Naâș) hors de la cellule et deux ions potassium (Kâș) dans la cellule
à l'aide de l'ATP comme source d'énergie. Cette pompe est présente dans
presque toutes les cellules animales. Elle est essentielle pour maintenir
un potentiel de membrane négatif, pour réguler le volume cellulaire et
pour créer un gradient ionique qui est utilisé pour d'autres processus
de transport.
âą La pompe calcium
ATPase (CaÂČâș-ATPase) transporte les ions calcium (CaÂČâș)
hors de la cellule ou dans les organites cellulaires, en utilisant Ă©galement
également l'ATP. Elle est importante pour la régulation du calcium intracellulaire,
qui est impliqué dans de nombreuses fonctions cellulaires, telles que
la contraction musculaire, la signalisation
cellulaire et la sécrétion.
âą La pompe Ă
proton (Hâș-ATPase) transporte les ions hydrogĂšne
(protons, Hâș) Ă travers la membrane cellulaire, contribuant ainsi au
maintien du pH intracellulaire et à la régulation de l'acidité cellulaire.
Par exemple, la pompe à protons présente dans les membranes des lysosomes
est essentielle pour maintenir l'acidité nécessaire à la dégradation
des déchets cellulaires.
âą La pompe Ă
chlorure (Clâ»-ATPase) transporte les ions chlorure
(Clâ») Ă travers la membrane cellulaire, contribuant ainsi Ă la rĂ©gulation
du volume cellulaire, de l'Ă©quilibre osmotique et du pH intracellulaire.
âą La pompe Ă
cations divalents (CaÂČâș, MgÂČâș-ATPase) transporte les ions divalents
tels que le calcium (CaÂČâș) ou le magnĂ©sium
(MgÂČâș) Ă travers la membrane cellulaire.
Endocytose.
L'endocytose est un type de transport
actif par lequel une cellule peut absorber des particules de dimensions
plus importantes, telles que des macromolécules, des parties de cellules
et mĂȘme des cellules entiĂšres.
Il existe différentes variantes d'endocytose,
mais toutes partagent une caractéristique commune : la membrane plasmique
de la cellule s'invagine, formant une poche autour de la particule cible.
La poche se pince, ce qui fait que la particule est enfermée dans une
vacuole nouvellement créée et qui est formée à partir de la membrane
plasmique. Puis la vacuole s'ouvre de l'autre cÎté de la membrane plasmique.
-
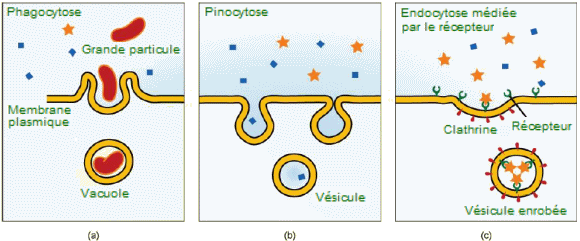
| Trois
variantes de l'endocytose. - (a) Dans la phagocytose, la membrane cellulaire
entoure la particule et s'invagine pour former une vacuole intracellulaire.
(b) Dans la pinocytose, la membrane cellulaire entoure un petit volume
de liquide et s'invagine, formant une vésicule. (c) Dans l'endocytose
médiée par les récepteurs, l'absorption de substances par la cellule
est ciblée sur un seul type de substance qui se lie au récepteur sur
la membrane cellulaire externe. (crédit : Mariana
Ruiz Villarreal). |
âą
La phagocytose. - C'est le processus par lequel de grosses particules,
telles que des cellules, sont absorbées par une cellule. La phagocytose
n'est observée que chez certains organismes unicellulaires tels que l'Amibe
(Les Protozoaires)
et dans les cellules animales telles que les leucocytes; ces derniers sont
des cellules sanguines responsables de la défense de l'organisme : les
leucocytes phagocytent les bactéries pathogÚnes ou nuisibles qui ont
pénétré dans l'organisme. Ainsi, un leucocyte appelé macrophage émet-il
d'abord des extensions de sa membrane appelées pseudopodes. Les
pseudopodes entourent les bactéries reconnues comme une menace, les extrémités
fusionnent pour former une grande vésicule (vacuole alimentaire) qui englobe
la bactérie. La vacuole alimentaire fusionne avec un lysosome dont les
enzymes digĂšrent la bactĂ©ries. Ce mĂȘme processus est effectuĂ© par des
organismes unicellulaires pour obtenir des nutriments.
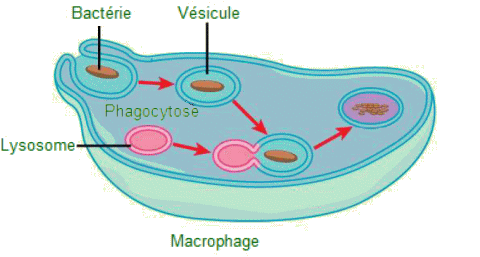
| Bactérie
potentiellement pathogÚne engloutie (phagocytée) par un macrophage.
- On observe les quatre Ă©tapes suivantes : 1) une portion de la membrane
plasmique du macrophage s'invagine ( = se replie) et engloutit un pathogĂšne;
2) la portion invaginée qui retient l'agent pathogÚne se détache de
la membrane plasmique et devient une vésicule; 3) la vésicule fusionne
avec un lysosome; 4) les enzymes hydrolytiques du lysosome détruisent
le pathogĂšne. |
âą La pinocytose.
- Cette autre variante de l'endocytose concerne presque toutes les cellules
eucaryotes, Ă la fois celles des organismes unicellulaires et celles des
animaux pluricellulaires, qui introduit constamment des fluides par pinocytose.
On a donné le nom de pinocytose (pino- = boire) à ce processus
Ă une Ă©poque oĂč l'on supposait que la cellule absorbait dĂ©libĂ©rĂ©ment
du liquide extracellulaire. En réalité, ce processus absorbe les solutés
du liquide extracellulaire dont la cellule a besoin .
âą L'endocytose
médiée par un récepteur. - Une variante ciblée de l'endocytose utilise
des protéines de liaison dans la membrane plasmique qui sont spécifiques
à certaines substances. Les particules se lient aux protéines et la membrane
plasmique s'invagine, amenant la substance et les protéines dans la cellule.
Si le passage à travers la membrane de la cible de l'endocytose médiée
par les récepteurs est inefficace, elle ne sera pas éliminé des fluides
tissulaires ou du sang. Au lieu de cela, elle restera dans ces fluides
oĂč sa concentration augmentera.
Certaines
maladies humaines sont causées par une défaillance de l'endocytose médiée
par les récepteurs. Par exemple, la forme de cholestérol
appelée lipoprotéine de basse densité ou LDL (également appelée
« mauvais » cholestérol) est éliminée du sang par endocytose médiée
par des récepteurs. Dans l'hypercholestérolémie familiale, une maladie
génétique humaine, les récepteurs LDL sont défectueux ou complÚtement
absents. Les personnes atteintes de cette maladie ont des taux de cholestérol
potentiellement mortels, car leurs cellules ne peuvent pas Ă©liminer ce
produit chimique de leur sang.
Exocytose.
L'exocytose est le mécanisme contraire
de l'endocytose. Son but est d'expulser un matériau de la cellule dans
le liquide extracellulaire. Ce peut ĂȘtre des dĂ©chets
issus de l'activité de la cellule ou des substances qu'elle sécrÚte
normalement, comme des hormones et les enzymes destinĂ©es Ă ĂȘtre utilisĂ©es
ailleurs dans l'organisme.
-
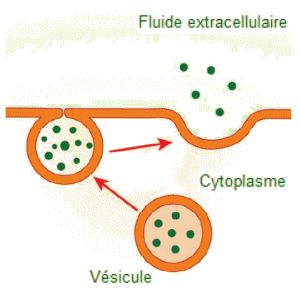
| Dans
l'exocytose, une vésicule migre vers la membrane plasmique, se
lie et libÚre son contenu à l'extérieur de la cellule.
(Crédit :Mariana Ruiz Villarreal). |
Le cytoplasme
Le cytoplasme comprend
deux parties :
âą
Le hialoplasme ou cytosol, qui est une solution aqueuse de sels, sucres,
acides aminés, protéines, acides gras, nucléotides et autres matiÚres.
De consistance gélatineuse, le cytosol est le milieu dans lequel baignent
les diverses structures intra-cellulaires : le noyau et les autres organites.
âą Le cytosquelette,
constitué d'un réseau de fibres et de tubules protéiques, forme un systÚme
de support intracellulaire.
De nombreuses réactions
biologiques se produisent dans le cytoplasme grùce aux enzymes (protéines)
qui catalysent ces réactions.
Les paragraphes qui
suivent passent en revue les organites communs Ă toutes les cellules eucaryotes,
puis ceux propres aux cellules animales et végétales. Le cytosquelette
et le noyau sont abordés ensuite.
Organites présents
dans toutes les cellules eucaryotes
Ribosomes.
De 15 Ă 30 nanomĂštres
de diamĂštre environ, les ribosomes sont les organites cellulaires les
plus nombreux. une cellule peut en contenir jusqu'Ă un demi-million. Lorsqu'ils
sont vus au microscope Ă©lectronique, les ribosomes libres apparaissent
sous forme de minuscules granules sombres isolés ou en amas (polyribosomes)
flottant librement dans le cytoplasme. Selon le type et la fonction de
la cellule concernĂ©e, les ribosomes peuvent aussi ĂȘtre attachĂ©s au cĂŽtĂ©
cytoplasmique de la membrane plasmique ou au cÎté
cytoplasmique du réticulum endoplasmique.
La microscopie Ă©lectronique
a montré que les ribosomes ne possÚdent pas de membrane et se composent
de deux sous-unités, une grande et une petite, chacune composée d'ARN
ribosomique (ARNr) et d'un grand nombre de protéines. Lorsque les deux
sous-unités se rejoignent, elles forment une machine moléculaire complexe,
dont les enzymes facilitent la synthÚse des protéines dans la
cellule.
Parce que la synthÚse des protéines est
essentielle pour toutes les cellules, les ribosomes se trouvent dans pratiquement
toutes les cellules, bien qu'ils soient plus petits dans les cellules procaryotes.
Ils sont particuliĂšrement abondants dans les globules rouges immatures
oĂč ils synthĂ©tisent l'hĂ©moglobine, la protĂ©ine impliquĂ©e dans le transport
de l'oxygĂšne dans tout le corps.
Mitochondries.
Les mitochondries sont souvent qualifiées
de « centrales énergétiques » de la cellule car elles sont responsables
de la fabrication de l'adénosine triphosphate (ATP), la principale molécule
porteuse d'Ă©nergie de la cellule. La formation d'ATP
à partir de la dégradation du glucose est connue sous le nom de respiration
cellulaire. Ce processus délivre l'énergie dont la cellule a besoin
pour mener à bien ses activités (synthÚse de substances, transport actif,
division cellulaire, mouvement des cils et des flagelles, etc.). On notera
ici que les cellules musculaires ont une concentration trÚs élevée de
mitochondries car les cellules musculaires ont besoin de beaucoup d'Ă©nergie
pour se contracter. Si une cellule ne reçoit pas d'énergie, non seulement
elle ne fonctionne pas, mais elle se désintÚgre également en peu de
temps.
Typiquement de la taille d'une bactérie,
les mitochondries affectent des formes trĂšs variĂ©es, elles peuvent ĂȘtre
cylindriques, sphériques, avec l'apparence d'une pomme de terre, mais
le plus souvent ovales. Elles possĂšdent une double membrane et ont
leurs propres ribosomes et ADN. Chaque membrane est une bicouche phospholipidique
incrustée de protéines. La couche interne a des plis appelés
crĂȘtes,
qui augmentent la surface de la membrane interne. La zone entourée par
les plis s'appelle la matrice mitochondriale; elle contient un liquide
trĂšs visqueux. Les crĂȘtes et la matrice ont des rĂŽles diffĂ©rents dans
la respiration cellulaire.
-
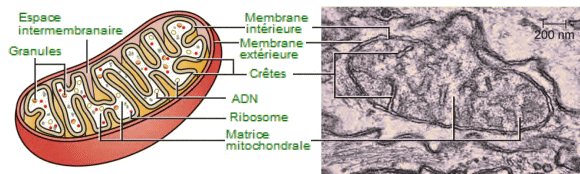
| Mitochondrie.
- Chaque mitochondrie est entourée
de deux membranes, dont l'intĂ©rieur est largement pliĂ© en crĂȘtes et
est le site de l'espace intermembranaire. La matrice mitochondriale contient
l'ADN mitochondrial, les ribosomes et les enzymes métaboliques. La micrographie
Ă©lectronique Ă transmission d'une mitochondrie, Ă droite, montre les
deux membranes, les crĂȘtes et la matrice mitochondriale.
(Micrographie : Matthew Britton). |
Peroxysomes.
Les peroxysomes sont de petits organites
ronds entourés de membranes simples. Ils effectuent des réactions d'oxydation
qui décomposent les acides gras et les acides aminés. Ils détoxifient
également de nombreux poisons qui peuvent pénétrer dans le corps (l'alcool,
par exemple, est détoxifié par les peroxysomes des cellules hépatiques).
Beaucoup de ces réactions d'oxydation libÚrent du peroxyde d'hydrogÚne,
qui pourrait endommager les composants cellulaires à l'extérieur de l'organite;
cependant, lorsque ces réactions se limitent aux peroxysomes, les enzymes
décomposent le peroxyde d'hydrogÚne en oxygÚne et en eau en toute sécurité.
Les cellules végétales possÚdent des
peroxysomes spécialisés, less glyoxysomes, qui sont responsables
de la conversion des graisses stockées en sucres. Les cellules végétales
contiennent de nombreux types de peroxysomes différents qui jouent un
rÎle dans le métabolisme, la défense des pathogÚnes et la réponse
au stress, pour n'en citer que quelques-uns.
RĂ©ticulum endoplasmique.
Le réticulum endoplasmique,
découvert en 1945 par Keith Porter, est une série de tubules membranaires
interconnectés qui modifient collectivement les protéines
et synthétisent les lipides. Ces deux fonctions
sont exécutées dans des zones distinctes du réticulum endoplasmique
: respectivement le réticulum endoplasmique rugueux,
et
le réticulum endoplasmique lisse.
La partie creuse des tubules s'appelle
la lumen (ou lumiĂšre) ou l'espace cisternal. La membrane
du réticulum endoplasmique qui est une bicouche phospholipidique incrustée
de protéines, est continue avec l'enveloppe nucléaire.
RĂ©ticulum
endoplasmique rugueux (ergatoplasme).
Le réticulum endoplasmique rugueux ou
granuleux est ainsi nommé parce que les ribosomes attachés à sa surface
cytoplasmique lui donnent un aspect clouté lorsqu'ils sont vus au microscope
Ă©lectronique.
Les ribosomes synthétisent les protéines
lorsqu'ils sont attachés au réticulum endoplasmique, ce qui entraßne
le transfert de leurs protéines nouvellement synthétisées dans la lumiÚre
du rĂ©ticulum endoplasmique rugueux oĂč ils subissent des modifications
telles que le pliage ou l'ajout de sucres. Le
réticulum endoplasmique rugueux fabrique également des phospholipides
pour les membranes cellulaires.
Si les phospholipides ou les protéines
modifiées ne sont pas destinés à rester dans le réticulum endoplasmique
rugueux, ils seront enrobés dans des vésicules et transportés du réticulum
endoplasmique rugueux par bourgeonnement Ă partir de la membrane. Ătant
donné que le réticulum endoplasmique rugueux est engagé dans la modification
des protéines qui seront sécrétées par la cellule, il est abondant
dans les cellules qui sécrÚtent des protéines, telles que le foie.
RĂ©ticulum
endoplasmique lisse.
Le réticulum endoplasmique lisse est
dans la continuité du réticulum endoplasmique rugueux mais a peu ou pas
de ribosomes sur sa surface cytoplasmique. Le réticulum
endoplasmique lisse synthétise des glucides, des lipides (graisses, phospholipides
et hormones stéroïdes); il joue aussi un rÎle dans la désintoxication
des médicaments et des poisons dans le métabolisme de l'alcool et le
stockage des ions calcium.
VĂ©sicules et
vacuoles.
Presque toutes les
cellules contiennent des vésicules et des vacuoles; ces organites sont
des sacs liés à la membrane plasmique qui interviennent lors du stockage
et du transport. Leur contenu dĂ©pend de la cellule oĂč elles se trouvent.
Les vacuoles
sont un peu plus grandes que les vésicules et la membrane d'une vacuole
ne fusionne pas avec les membranes d'autres composants cellulaires. Les
vésicules peuvent fusionner avec d'autres membranes du systÚme cellulaire.
Les enzymes présentes dans les vacuoles des plantes
peuvent briser les macromolécules.
Ajoutons que la plupart
des Protozoaires ont des vacuoles digestives
avec des aliments en cours de digestion, en plus des vacuoles contractiles
qui expulsent à l'extérieur de la cellule et de l'excÚs d'eau. Par exemple,
dans un Protozoaire d'eau douce comme la Paramécie, l'eau pénÚtre constamment
dans la cellule par osmose. Si l'excÚs d'eau n'était pas éliminé par
les vacuoles contractiles, cet organisme exploserait.
Appareil de Golgi.
Nous avons dit que les vésicules peuvent
bourgeonner du réticulum endoplasmique et transporter leur contenu ailleurs.
Avant d'atteindre leur destination finale, les lipides ou les protéines
inclus dans les vĂ©sicules de transport doivent ĂȘtre triĂ©s, empaquetĂ©s
et étiquetés afin qu'ils se retrouvent au bon endroit. Le tri, le marquage,
le conditionnement et la distribution des lipides et des protéines ont
lieu dans l'appareil de Golgi (également appelé corps de Golgi), du nom
de son découvreur en 1898, l'histologiste Camilo Golgi, qui lui l'appelait
l'appareil
à réseau interne.
L'appareil de Golgi correspond à une série
de sacs membraneux aplatis appelés saccules membranaires. Un empilement
de tels saccules correspond Ă un dictyosome. On y distingue une face
cis, ou face de réception cÎté réticulum endoplasmique, et
une face trans, ou de libération, du cÎté opposé au réticulum
endoplasmique, vers la membrane cellulaire.
-
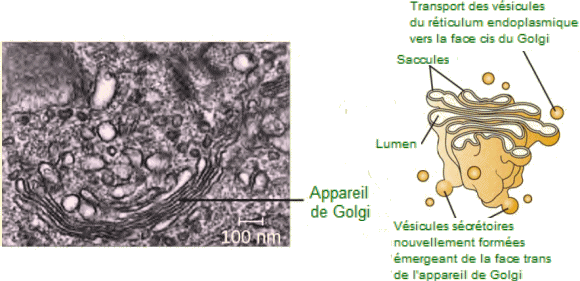
-
| L'appareil
de Golgi sur cette micrographie Ă©lectronique Ă transmission d'un
globule blanc est visibl sous la forme d'un empilement d'anneaux aplatis
semi-circulaires dans la partie inférieure de l' image. Plusieurs vésicules
peuvent ĂȘtre vues prĂšs de l'appareil de Golgi. L'illustration (Ă droite)
montre les disques empilés en forme de coupe et plusieurs vésicules de
transport. L'appareil de Golgi modifie les lipides et les protéines, produisant
des glycolipides et des glycoprotéines, respectivement, qui sont généralement
insérés dans la membrane plasmique.(Photo :
Louisa Howard). |
Les vésicules de transport qui se forment
à partir du réticulum endoplasmique se dirigent vers la face de réception,
fusionnent avec elle et vident leur contenu dans la lumiĂšre d'un dictyosome
de l'appareil de Golgi.
La progression continue à l'intérieur
de l'appareil de Golgi, et, Ă mesure qu'ils avancent,
les protéines et les lipides subissent d'autres modifications. La modification
la plus fréquente est l'ajout de chaßnes courtes de molécules de sucre
pour fabriquer des glycoprotéines et des glycolipides. Les protéines
et lipides nouvellement modifiés sont ensuite marqués avec de petits
groupes moléculaires (groupes phosphate ou autres petites molécules)
pour leur permettre d'ĂȘtre acheminĂ©s vers leurs destinations appropriĂ©es.
Pour finir, les protéines modifiées et
marquées sont empaquetées dans des vésicules sécrétoires qui bourgeonnent
Ă partir de la face trans du dictyosome. Alors
que certaines de ces vésicules déposent leur contenu dans d'autres parties
cellulaires oĂč elles seront utilisĂ©es, d'autres vĂ©sicules sĂ©crĂ©toires
fusionnent avec la membrane plasmique et libÚrent leur contenu à l'extérieur
de la cellule.
La quantité de dictyosomes dans l'appareil
de Golgi des différents types de cellules de la fonction des cellules.
Les cellules qui exercent une grande activité sécrétoire (comme les
cellules des glandes salivaires qui sécrÚtent
des enzymes digestives ou les cellules du systÚme immunitaire qui sécrÚtent
des anticorps) ont un nombre abondant de telles structures.
Dans les cellules végétales, l'appareil
de Golgi a un rÎle supplémentaire de synthÚse de polysaccharides, dont
certains (la cellulose) sont incorporés dans la paroi cellulaire et certains
sont utilisés dans d'autres parties de la cellule.
Organites présents
dans les cellules animales seules (sauf exceptions)
Centrosome et
centrioles
Le centrosome, que l'on rencontre à proximité
du noyau des cellules animales (ainsi que des cellules
de certaines Algues et de Champignons) est un organite composé d'une paire
de corps cylindriques creux, perpendiculaires l'un avec l'autre, et appelés
centrioles.
Chaque centriole est un cylindre de neuf triplets de microtubules.
Le centrosome se réplique avant qu'une
cellule ne se divise, et les centrioles jouent un rĂŽle en tirant les chromosomes
dupliqués aux extrémités opposées de la cellule en cours de division.
Cependant, la fonction exacte des centrioles dans la division cellulaire
n'est pas claire, car les cellules dont les centrioles ont disparu peuvent
toujours se diviser et les cellules végétales, qui manquent de centrioles,
sont capables de division cellulaire.
Les centrioles donnent
également naissance aux cils et aux flagelles. Dans ce cas, étant situés
Ă la base de ces structures motrices, ils prenent le nom de corps basaux.
-
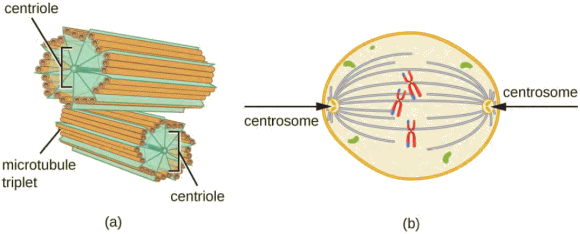
| (a)
Un
centrosome est composé de deux centrioles positionnés à angle droit
l'un par rapport à l'autre. Chaque centriole est composé de neuf triplets
de microtubules maintenus ensemble par des protéines accessoires. (b)
Dans les cellules animales, les centrosomes (flĂšches) servent de centres
d'organisation des microtubules du fuseau mitotique pendant la mitose. |
Flagelles et cils.
Les flagelles et les cils sont des structures
constituées de microtubules, en forme de cheveux, et qui s'étendent hors
de la cellule Ă partir de la membrane plasmique. Les flagelles sont longues,
les cils sont courts.
Les flagelles sont utilisées pour déplacer
une cellule entiĂšre (par exemple, le spermatozoĂŻde, Euglena). Les cellules
qui en possĂšdent n'ont qu'une ou un petit nombre de flagelles. Note :
les flagelles peuvent aussi se rencontrer dans des cellules non animales
: chez les Eucariotes, Chlamydomonas, une algue verte, en possĂšde
deux; chez les Procaryotes,Helicobacter pylori, une bactérie, en possÚde
une dizaine, etc.
Lorqu'une cellule possĂšde des cils, ceux-ci
sont extrĂȘment nombreux et sont prĂ©sents sur toute la surface de la membrane
plasmique (comme la Paramécie qui possÚde jusqu'à 17 000 cils). Par
leurs mouvements coordonnés les cils permettent le déplacement
des cellules entiÚres, mais également le déplacement de substances diverses
le long de la surface externe de la cellule. Il en est ainsi des cils des
cellules tapissant les trompes de Fallope
qui déplacent l'ovule vers l'utérus,
ou des cils tapissant les cellules des voies
respiratoires qui déplacent les particules que le mucus a piégé
vers la gorge.
Certains Procaryotes
ont aussi des flagelles, des pili (plus ou moins analogues au cils)
ou des fimbriae (franges, pili de petite taille). La plupart des
pili sont utilisés pour échanger du matériel génétique lors d'un type
de reproduction appelé conjugaison. Les flagelles, comme dans le
cas des Eucaryotes, sont utilisées pour la locomotion.
-
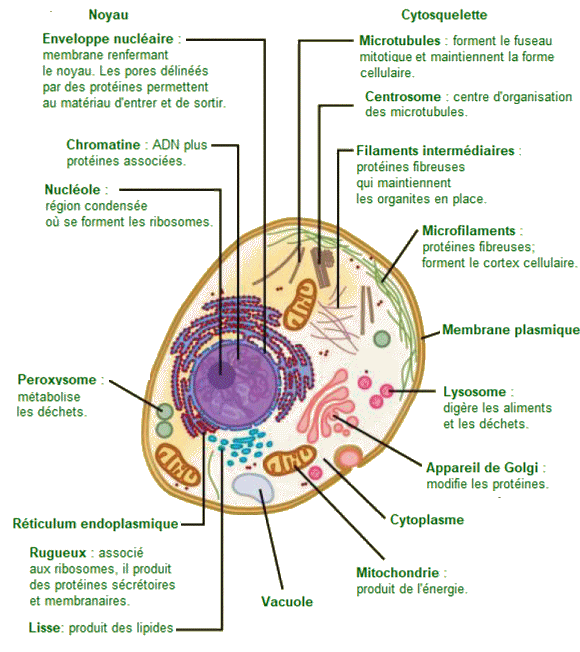
Organites
d'une cellule animale typique.
Lysosomes.
Les lysosomes,
découverts en 1949 par Christian de Duve, sont des vésicules particuliÚres,
d'une taille de l'ordre du micromÚtre, dotées d'une fonction de dégradation
et d'élimination des déchets cellulaires, et qui contiennent pour cela
d'un grand nombre (on en a dénombré une cinquantaine) d'enzymes digestives
(hydrolases). Ces enzymes aident à la dégradation des protéines, des
polysaccharides, des lipides, des acides nuclĂ©iques et mĂȘme des organites
usés.
Chez les Eucaryotes unicellulaires, les
lysosomes ont un rÎle important dans la digestion des aliments ingérés
et dans le recyclage des organites. Les enzymes qu'ils renferment sont
actives à un pH beaucoup plus bas (plus acide) que celles situées dans
le cytoplasme. De nombreuses réactions qui ont lieu dans le cytoplasme
ne pourraient pas se produire Ă un pH bas, d'oĂč l'avantage de compartimenter
la cellule eucaryote pour disposer de lieux, comme l'intérieur des lysosomes,
oĂč certaines rĂ©actions chimiques sont possibles alors qu'elles ne le
sont pas ailleurs.
Les lysosomes utilisent Ă©galement leurs
enzymes hydrolytiques pour détruire les organismes pathogÚnes qui pourraient
pénétrer dans la cellule. Un bon exemple de cela est donné par un groupe
de globules blancs appelés macrophages, qui font partie du systÚme immunitaire
: ors d'un processus connu sous le nom de phagocytose (voir plus
haut).
(Les lysosomes, qui sont des organites
propres aux cellules animales ont leur équivalent chez les végétaux
: ce sont les phytolysosomes, intégrés dans la vacuole centrale,
mais ceux-ci ont des fonctions moins Ă©tendues que les lysosomes ).
Organites présents
dans les cellules végétales seules
Paroi cellulaire.
La paroi cellulaire
est un revĂȘtement rigide qui protĂšge la cellule, lui fournit un support
structurel et lui donne sa forme. C'est aussi cette rigidité qui permet
aux plantes de se desser au-dessus du sol. Outres les plantes, les algues,
les champignons et les bactéries ont une paroi cellulaire, qui enveloppe
la membrane plasmique.
La principale molécule
organique de la paroi cellulaire végétale est un polysaccharide composé
de longues chaßnes droites d'unités de glucose, la cellulose. On
rencontre aussi dans la paroi cellulaire de la pectine, une substance gélatineuse
qui agit comme ciment entre les cellules.
On a vu plus haut
que la paroi cellulaire comporte de minuscules trous, les plasmodesmes,
Ă travers lesquels un contact s'Ă©tablit entre une cellule et une autre,
permettant aussi un Ă©change de substances entre elles. Mais contrairement
à la membrane plasmique, la paroi cellulaire, perméable à la plupart
des molécules, ne contrÎle pas le passage des matériaux dans et hors
de la cellule.
Les Bactéries,
à l'exception des Microplasmes (les plus petites Bactéries) ont une paroi
cellulaire en peptidoglycane (contrairement aux Archées et aux Plantes),
composée de sucres et d'acides aminés, et beaucoup ont une capsule de
polysaccharide. Dans leur cas, comme dans celui des Plantes,
la paroi cellulaire agit comme une couche de protection supplémentaire,
aide la cellule Ă conserver sa forme et empĂȘche la dĂ©shydratation. La
capsule permet Ă la cellule de se fixer aux surfaces de son environnement.
-
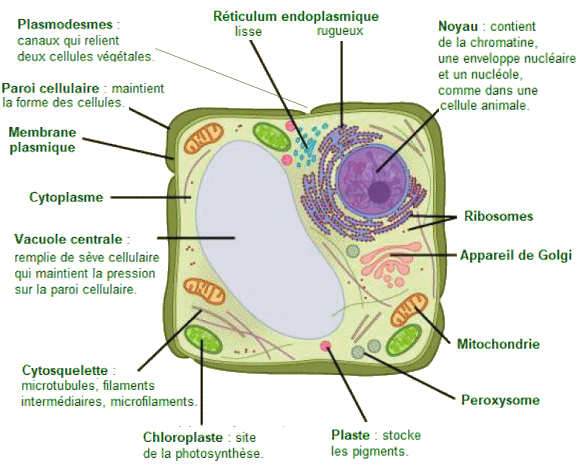
Cellule
végétale typique.
Les plastes. -
Les
plastes ou plastides sont des organites que l'on ne trouve que dans
les cellules des Plantes et des Algues. Ces organites, entourés de deux
membranes, sont de trois types : les chloroplastes, les chromoplastes et
les leucoplastes.
Chloroplastes.
D'une taille généralement
comprise entre 3 et 10 micromĂštres de long, les chloroplastes sont les
plastes les plus courants et ceux qui ont la plus grande importance biologique,
car la vie sur notre planÚte dépend de leur fonction, qui est le processus
de photosynthĂšse. Comme les mitochondries, les chloroplastes (chloro-
= vert) ont Ă©galement leur propre ADN et leurs propres ribosomes. D'ailleurs,
selon la théorie endosymbiotique de l'origine de la cellule eucaryote,
la présence de chloroplastes, comme celle des mitochondries dans les cellules
auraient une origine similaire. Il s'agirait d'anciennes bactéries absorbées
et intégrées dans le métabolisme des cellules eucaryotes : les chloroplastes
dériveraient de cyanobactéries,
tandis que les mitochondries dériveraient de bactéries respirantes
Dans la photosynthĂšse,
le dioxyde de carbone, l'eau et l'énergie lumineuse sont utilisés pour
fabriquer du glucose et de l'oxygÚne. C'est la principale différence
entre les Plantes et les Animaux : les Plantes (autotrophes) sont capables
de fabriquer leur propre nourriture, comme le glucose, tandis que les Animaux
(hétérotrophes) doivent compter sur d'autres organismes pour leurs composés
organiques ou leur source de nourriture.
Les chloroplastes
sont entourés d'une membrane externe et d'une membrane interne; dans l'espace
clos par la membrane interne d'un chloroplaste se trouve un ensemble de
sacs membranaires appelés thylakoïdes,
qui sont interconnectés et empilés et remplis de liquide. Chaque pile
de thylakoĂŻdes prend le nom de granum (pluriel,
grana).
Le fluide enfermé par la membrane interne et entourant les grana étant
quant à lui nommé stroma.
Les grana contiennent un pigment vert,
la chlorophylle, qui donne leur couleur aux plantes et surtout qui capture
l'Ă©nergie de la lumiĂšre solaire pour la photosynthĂšse.
-
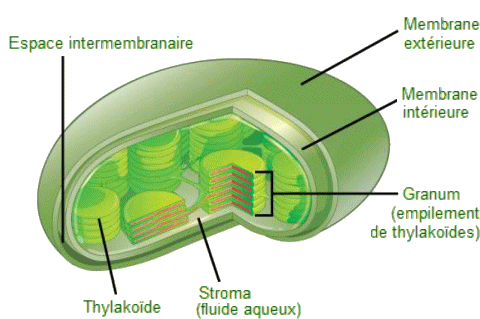
Chloroplaste.
- Ce diagramme simplifié d'un chloroplaste montre la membrane externe,
la membrane interne, les thylakoĂŻdes, les grana et le stroma. |
Comme les cellules végétales, les Protistes
photosynthétiques ont également des chloroplastes. Certaines Bactéries
effectuent Ă©galement la photosynthĂšse, mais elles n'ont pas de chloroplastes.
Leurs pigments photosynthétiques sont situés dans la membrane thylakoïde
Ă l'intĂ©rieur de la cellule elle-mĂȘme.
En plus de la chlorophylle,
certains chloroplastes contiennent des pigments de diverses couleurs. Ordinairement,
ils sont masqués par l'abondance de la chlorophylle, mais peuvent donner
leur couleur aux cellules et aux organismes qu'elles composent, si la teneur
en chlorophylle dimine.
Chromoplastes.
Les chromoplastes
(chromo- = couleur) sont les plastes responsables des couleurs rouges,
jaunes ou oranges des fruits, des légumes, des
fleurs
et des feuilles des arbres en automne. Ces couleurs
sont dues aux pigments appelés
caroténoïdes, qui sont divisés
en deux groupes : les carotĂšnes de couleurs orange et rouge et
les xanthophylles de couleur jaune.
Les chromoplastes
peuvent ĂȘtre dĂ©rivĂ©s des chloroplastes lorsqu'ils perdent de la chlorophylle.
Le changement de couleur des fruits verts lors
de la maturation, dans de nombreuses Plantes, est dĂ» Ă la conversion
des chloroplastes en chromoplastes. Par exemple, lorsqu'une tomate mûrit,
la chlorophylle est détruite, révélant la couleur rouge, initialement
masquée par la chlorophylle. D'autres pigments qui donnent de la couleur
aux fleurs et Ă d'autres parties des plantes se trouvent dissous dans
des vacuoles. Ces pigments sont des
anthocyanes, dont la gamme de
couleurs va du bleu ou du violet au rouge.
Leucoplastes.
Les leucoplastes
(leuco - = blanc) sont incolores, leur fonction est de stocker l'amidon
et d'autres substances telles que les protéines et les lipides. Ils sont
trĂšs nombreux dans les racines (navets) ou
dans les tubercules (pommes de terre). Ils
peuvent Ă©galement ĂȘtre trouvĂ©s dans d'autres parties des plantes : dans
les graines, ou dans l'Ă©piderme des feuilles,
des tiges et des fleurs.
La vacuole centrale.
Chaque cellule végétale
possĂšde une grande vacuole centrale qui occupe la majeure partie de la
cellule (couramment jusqu'aux trois quarts de son volume). Une fonction
de cette vacuole est de stocker diverses substances de réserve telles
que l'amidon, des protéines, des graisses et des pigments, en particulier
dans les cellules germinales en développement. Mais surtout, la vacuole
centrale joue un rÎle clé dans la régulation de la concentration
en eau de la cellule, en cas de changements dans
les conditions environnementales.
-
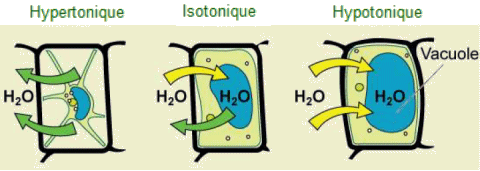
| La
pression de turgescence à l'intérieur d'une cellule végétale dépend
de la tonicité de la solution dans laquelle elle est baignée.
(Crédit :Mariana Ruiz Villarreal). |
Lorsqu'elle est gonflée,
la vacuole centrale exerce une pression contre la paroi cellulaire, procurant
ainsi à la cellule un degré élevé de fermeté ou de turgescence, ce
qui permet aux feuilles, aux fleurs et aux tiges tendres des plantes de
rester fermes. Ainsi, si l'on oublie d'arroser une plante pendant quelques
jours, elle flétrit. En effet, à mesure que la concentration en
eau dans le sol devient inférieure à la concentration en eau dans la
plante, l'eau quitte la vacuole centrale et le cytoplasme de cellules pour
pénétrer dans le sol. Lorsque la vacuole centrale se vide et se contracte,
elle laisse la paroi cellulaire sans support. C'est cette perte de support
des parois cellulaires qui donne l'aspect flétri aux plantes manquant
d'eau.
Le cytosquelette
DĂ©couvert dans les
années 1970 grùce au microscope électronique, le cytosquelette, qui
s'étend en forme de réseau dans tout le cytoplasme, représente l'armature
de la cellule. Les fonctions du cytosquelette sont multiples : il soutient
et donne sa forme à la cellule; il maintient et déplace les organites;
il participe au mouvement de la cellule par les déformations qu'il imprime
Ă la cellule (mouvement amiboĂŻde).
Il existe trois types de fibres dans le
cytosquelette : les microfilaments, également appelés filaments d'actine,
les filaments intermédiaires et les microtubules.
-
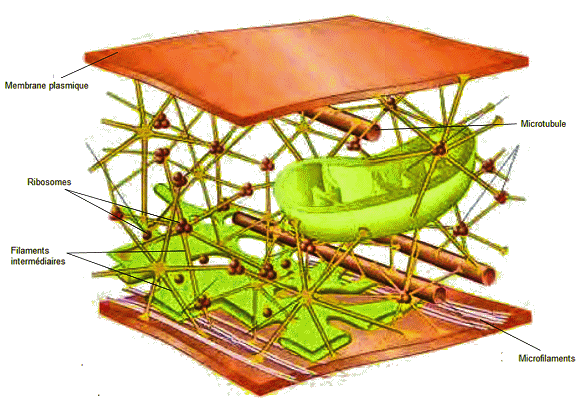
Le
cytosquelette.
Source
: Université autonome du Sinaloa.
Les microfilaments.
Les microfilaments, principalement composés
des protéines actine et myosine, sont les fibres les plus minces du cytosquelette
et interviennent dans les composants cellulaires en mouvement, par exemple,
pendant la division cellulaire. Ils maintiennent Ă©galement la structure
des microvillosités, et le pliage étendu de la membrane plasmique que
l'on trouve dans les cellules dédiées à l'absorption. Ces composants
sont Ă©galement communs dans les cellules musculaires et sont responsables
de leur contraction.
Les filaments
intermédiaires.
Les filaments intermédiaires, dont les
constituants sont des protéines différentes selon les cellules, sont
de diamÚtre intermédiaire et ont des fonctions structurelles, telles
que le maintien de la forme de la cellule et l'ancrage des organites. La
kératine, par exemple, qui renforce les cheveux
et les ongles, est le composant d'un type de filament
intermédiaire.
Les microtubules.
Les microtubules,
constitués de la protéine tubuline, sont les fibres les plus épaisses
du cytosquelette. Ce sont des tubes creux qui peuvent se dissoudre et se
reformer rapidement. Les microtubules guident le mouvement des organites
et sont les structures (fuseau mitotique) qui
attirent les chromosomes vers leurs pĂŽles pendant la division cellulaire.
Ils sont Ă©galement les composants structurels des flagelles et des cils.
Les flagelles et les cils sont organisĂ©s de la mĂȘme façon : neuf
microtubules doubles forment un cercle à l'extérieur, tandis que deux
autres microtubules sont au centre.
Le noyau
En rÚgle générale, le noyau est l'organite
le plus important dans une cellule eucaryote. Le noyau abrite l'ADN (matériel
génétique) de la cellule et dirige la synthÚse des ribosomes et des
protéines. La plupart des cellules ont un seul noyau,
mais certaines, issues d'une fusion de cellules, en ont deux (c'est souvent
le cas des cellules du foie) ou mĂȘme bien davantage,
comme dans le cas des cellules musculaires striées qui composent les muscles
(plusieurs centaines de noyaux). Il existe aussi de cellules, chez les
Eucaryotes, qui ne possĂšdent pas de noyau ou qui l'on perdu (ex. : les
globules rouges). En général les cellules anuclées ne survivent pas
longtemps.
Le noyau, d'un diamĂšre
de l'ordre de 5 ”m, se situe le plus souvent prÚs du centre de la cellule,
mais il peut ĂȘtre trouvĂ© n'importe oĂč dans la cellule; sa forme peut
ĂȘtre sphĂ©rique ou ovale. Il est composĂ© de quatre parties : l'enveloppe
nucléaire, le nucléoplasme, le nucléole et la chromatine.
-
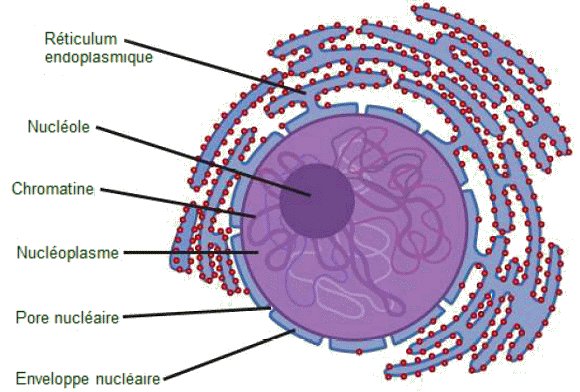
| Le
noyau cellulaire. - Le noyau stocke la chromatine (ADN plus protéines)
dans une substance semblable à un gel appelée nucléoplasme. Le nucléole
est une rĂ©gion condensĂ©e de la chromatine oĂč se produit la synthĂšse
des ribosomes. La limite du noyau s'appelle l'enveloppe nucléaire. Celle-ci
se compose de deux bicouches (membranes) phospholipidiques: une membrane
externe et une membrane interne, contrairement Ă la membrane plasmique,
qui se compose d'une seule bicouche phospholipidique. La membrane nucléaire
se prolonge sanns discontinuité par le réticulum endoplasmique. Les pores
nucléaires permettent aux substances d'entrer et de sortir du noyau. (Crédit
: NIGMS, NIH). |
L'enveloppe nucléaire.
L'enveloppe nucléaire
ou membrane nucléaire constitue la séparation entre le cytoplasme et
le noyau. Elle se compose de deux membranes, séparées par un espace de
l'ordre de 10 Ă 50 nm. En deux nombreux points (plusieurs milliers), ces
membranes se rejoignent pour former de petites ouvertures circulaires,
appelĂ©es pores nuclĂ©aires. Ce sont les passages permettant Ă
la matiÚre d'entrer dans le noyau (ex. les nucléotides) ou d'en sortir
(ex. l'ARNm). L'enveloppe nucléaire se désassemble au moment de la division
cellulaire pour se recontituer dÚs la télophase.
Le nucléoplasme.
Le nucléoplasme
ou caryoplasme est une substance semi-liquide, riche en protéines. Retenu
dans le noyau par l'enveloppe nucléaire, on peut y distinguer, comme pour
le cytoplasme plusieurs composantes : la composante liquide (70 % Ă 90
% d'eau) prend le nom de nucléosol ou de
hyaloplasme nucléaire.
On y trouve en suspension la chromatine et les nucléoles, et en solution
de nombreux composés : enzymes et autres protéines, nucléotides triphosphates,
etc.
La chromosomes
et chromatine.
La chromatine et les chromosomes ont la
mĂȘme composition chimique : le matĂ©riau qui les constitue est appelĂ©
chromatide;
c'est une combinaison d'ADN (nucléofilament), de protéines histones et
d'autres protĂ©ines. La chromatide peut revĂȘtir deux aspects diffĂ©rents
en fonction des Ă©tapes de la vie d'une cellule : chromosomes ou chromatine,
donc.
Chez les cellules eucaryotes, les chromosomes,
apparaissent comme des structures linéaires. Chaque espÚce a un nombre
spécifique de chromosomes dans le noyau de ses cellules. Par exemple,
chez l'Humain, le nombre de chromosomes est de 46, tandis que chez les
Mouches des fruits, le nombre de chromosomes est de 8. Les chromosomes
ne sont visibles et se distinguent les uns des autres que lorsque la cellule
s'apprĂȘte Ă se diviser. Ce sont des structures
de de 5 à 20 ”m de long.
Lorsque la cellule est dans les phases
de croissance et d'entretien de son cycle de vie, les chromosomes ressemblent
à un tas de fils déroulés et embrouillés, formant des structures de
10 000 à 40 000 ”m de long. et ils ne sont plus discernables. C'est alors
qu'on parle de chromatine.
-
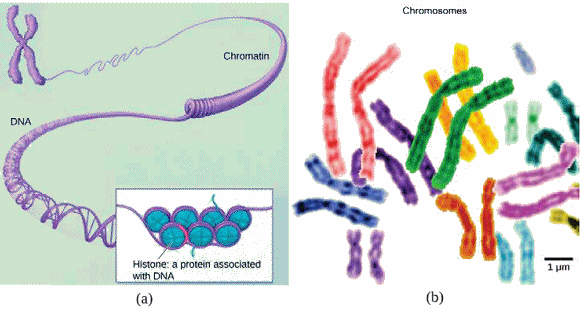
Chromatine
et chromosomes. - a) Différents niveaux d'organisation de la chromatine
(ADN
associé
à des histones). b) Chromosomes apariés. (Source
: NIH.).
Les nucléoles.
Nous avons dit que le noyau dirige la
synthĂšse des ribosomes. Certains chromosomes ont
des sections d'ADN qui codent pour l'ARN ribosomal. Une zone sphérique,
sans membrane qui la délimiterait, et de coloration sombre dans le noyau,
appelĂ©e nuclĂ©ole (il peut y en avoir plusieurs dans un mĂȘme noyau,
autour de régions chromosomiques appelées régions organisatrices nucléolaires),
agrÚge l'ARN ribosomal avec les protéines associées pour assembler les
sous-unités ribosomales qui sont ensuite transportées à travers
les pores nucléaires dans le cytoplasme. Lors de la division cellulaire,
quand la chromatine se condense en chromosome, la matiÚre des nucléoles
se disperse et on la retrouve ensuite recondensée dans les nouvelles cellules
filles. (D'aprĂšs des publications de la collection
Openstax
de l'Université Rice). |
|
